CHAPITRE 1: INTRODUCTION
Ce cours s'intéresse à l'étude des tissus, souvent appelée "histologie". Le mot tissu -tissage ou texture- fut introduit dans le langage de la biologie en 1801 par l'anatomiste et physiologiste Bichat, à partir d'observations à l'oeil nu de dissections d'organes. Par la suite, les tissus furent étudiés davantage à l'aide du microscope et le terme histologie fut introduit en 1839 par le microscopiste Schwann. C'est en histologie qu'on apprend le nom de tout ce qui peut être identifié au microscope, tant dans l'organisme végétal qu'animal. Ce cours se limite à l'étude du règne animal.
Les tissus sont formés par des cellules et ils forment eux-mêmes les organes. Ainsi, l'histologie comprend trois niveaux d'étude: l'étude des cellules ou cytologie, l'étude des tissus ou histologie proprement dite et l'étude des organes ou organologie. La cytologie fait l'objet d'un cours à part mais plusieurs des notions qui y ont été apprises serviront ici. Nous nous concentrerons sur l'étude des tissus de base, avec les cellules et le matériel intercellulaire qui les composent, dans un premier temps, et de l'organisation de ces tissus pour former les organes, dans un deuxième temps. Les séances de laboratoire porteront en plus sur l'aspect comparatif de ces tissus, surtout chez les vertébrés.
Il existe quatre tissus de base: épithélial, conjonctif, musculaire et nerveux, chacun ayant des subdivisions. Les tissus diffèrent les uns des autres parce que les cellules qui les constituent sont structuralement spécialisées pour accomplir des fonctions particulières. Ainsi, les quatre tissus de base ont été classés selon leur structure microscopique et leur fonction, non pas selon leur origine embryonnaire (voir 1.2). En effet, chez l'embryon on décrit trois feuillets tissulaires de base: l'ectoderme, le mésoderme et l'endoderme, desquels dérivent les tissus adultes. La notion de tissu fut d'une grande importance dans le développement de l'embryologie. Une fois l'origine embryonnaire des tissus déterminée, il devint plus facile de suivre leur organisation en organes (organogénèse) et leur croissance. De plus, la notion de tissu fut importante dans le développement de la pathologie.
1.1 METHODES DE BASE EN HISTOLOGIE
1.1.1 ETUDE IN VITRO
Il est possible d'isoler des cellules ou même des portions de tissus et organes, de les dissocier et de les maintenir dans des milieux de culture, qu'il devient possible d'étudier en détail par la microscopie et autres méthodes. En culture, les cellules peuvent, jusqu'à un certain point, se diviser, se mouvoir, sécréter et accomplir plusieurs des fonctions qu'elles accompliraient dans l'organisme. Elles peuvent parfois, dans les conditions appropriées, devenir plus spécialisées et exprimer des caractères nouveaux.
Cette technique permet d'effectuer des microchirurgies, comme transplanter le noyau d'une cellule dans le cytoplasme énucléé d'une autre, fusionner deux cellules provenant de tissus ou organismes différents, etc. L'avantage des cultures de tissus est de pouvoir observer et manipuler les cellules dans un environnement bien contrôlé, qui simule ou non celui de l'organisme, mais sans les influences complexes et variables qui les affectent dans l'organisme. Le désavantage principal réside justement dans la perte des interactions qui existent entre les cellules dans un tissu, dans un organe et dans l'organisme entier. Ce milieu demeure donc artificiel.
1.1.2 ETUDE DES CELLULES ET TISSUS EN COUPES HISTOLOGIQUES
Les coupes fines de tissus permettent d'étudier l'organisation structurale de populations cellulaires et même les modifications de cette organisation selon les conditions physiologiques ou pathologiques, avec l'âge, etc. Cette approche implique l'étude de cellules mortes, contrairement aux cultures, mais assure la préservation des relations structurales entre les cellules dans le tissu et l'organe. C'est la méthode des coupes histologiques que nous utiliserons dans ce cours pour l'examen des tissus et organes. L'obtention de coupes histologiques nécessite plusieurs manipulations dont il est important de comprendre les principes pour en apprécier les résultats d'après leurs valeurs, sans les surestimer.
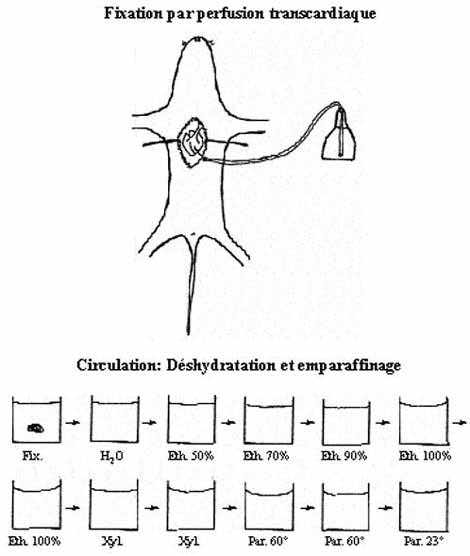
1.1.2.1 Fixation
Pour que les composantes des tissus soient préservées, l'organe doit être traité avec des produits chimiques qui en empêchent la détérioration; c'est la fixation. Les produits chimiques employés s'appellent fixateurs, préparés en solution aqueuse. La fixation s'effectue soit par immersion, en baignant dans le fixateur l'organe (entier ou tranché en pièces) disséqué de l'animal anesthésié (et euthanasié), soit en remplaçant par le fixateur le sang de l'animal anesthésié, par perfusion transcardiaque. Les fixateurs chimiques s'unissent aux protéines des tissus ou les dénaturent et les précipitent en remplaçant l'eau qui leur est associée. Les fixateurs inactivent les enzymes. Ils agissent comme antiseptiques en tuant les bactéries et autres microorganismes. Les fixateurs communément employés sont le formaldéhyde, le paraformaldéhyde et le glutaraldéhyde, seuls ou en combinaison avec d'autres produits. Certains fixateurs spéciaux peuvent être employés pour la préservation de composantes tissulaires spécifiques.
1.1.2.2 Enrobage
Il est difficile de couper un organe fixé en tranches suffisamment minces pour être observables au microscope. Puisqu'une chandelle, en réalité un bloc de cire, peut être tranchée très finement, l'idée est venue aux histologistes pionniers d'infiltrer de cire (paraffine) liquide les organes et de les laisser se solidifier. Cette méthode s'utilise encore et permet d'obtenir des coupes d'organes aussi minces que 5µm. Toutefois, comme la paraffine n'est pas miscible avec l'eau contenue dans l'organe et le fixateur, il faut déshydrater l'organe dans des bains successivement plus concentrés d'éthanol, puis d'éthanol absolu; c'est le processus de déshydratation. Puisque l'alcool n'est pas miscible avec la paraffine, l'organe infiltré d'alcool doit être baigné dans du dioxane ou du xylène avant d'être placé dans des bains de paraffine liquide. Dioxane et xylène sont miscibles tant dans l'alcool que dans la paraffine. Une fois l'enrobage achevé, c'est à dire que l'organe est infiltré et enrobé de paraffine et le dernier bain de paraffine est refroidi, on obtient un bloc qui sera facilement débité.
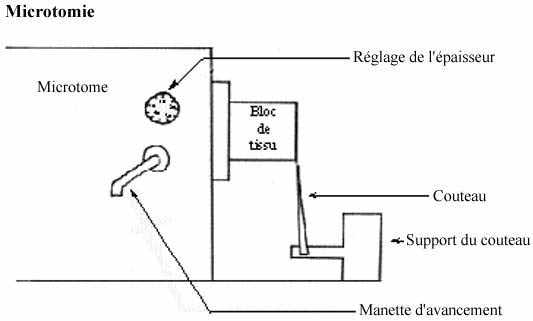
1.1.2.3 Confection des coupes (microtomie) et montage sur lame
Le bloc est placé sur la base d'un microtome, appareil muni d'un couteau et d'un mécanisme d'avancement de sa base, qui permet de trancher le bloc à l'épaisseur désirée. Pour des études histologiques routinières, l'épaisseur des coupes varie entre 10 et 20µm.
La méthode décrite ci-haut est la plus commune pour étudier l'histologie générale. Toutefois, il existe d'autres façons de préparer des coupes histologiques. Par exemple, l'organe, fixé ou non, peut être congelé dans de la glace sèche ou de l'azote liquide (qui servent de fixateur) et monté sans enrobage sur un microtome à congélation, ou cryotome. Ainsi congelé, l'organe est suffisamment dur pour être coupé finement. Cette technique présente l'avantage d'obtenir des résultats très rapidement, mais de qualité moindre. L'enrobage d'une pièce d'organe peut se faire avec du plastique ou des résines, plutôt qu'à la paraffine, lorsque des coupes de l'ordre de 1µm ou moins sont désirées.
Afin de pouvoir être observées au microscope, les coupes obtenues sont montées sur des lames de verre préalablement recouvertes de "colle" d'albumine ou de gélatine pour en assurer l'adhérence, puis sont séchées; c'est le montage sur lame.
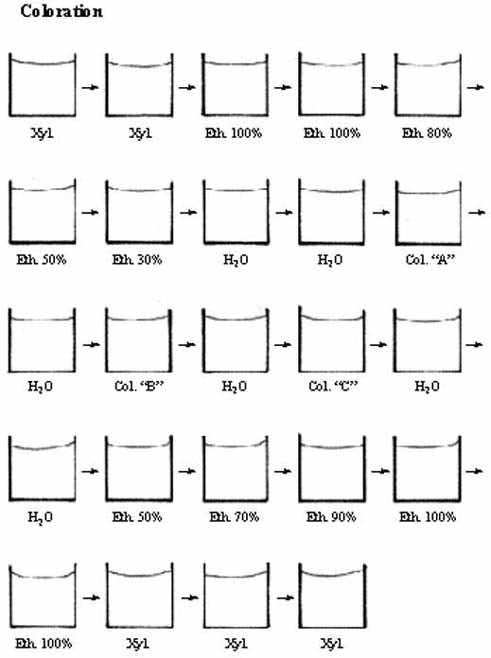
1.1.2.4 Coloration
Des coupes simplement montées sur lames s'avèrent presque toujours insuffisantes pour l'étude microscopique puisque le seul moyen de distinguer la structure d'un tissu est la différence entre les indices de réfraction de ses composantes. Or, les composantes cellulaires ont à peu près toutes la même densité optique. Toutefois, il est possible de colorer le tissu et d'en dévoiler la constitution microanatomique à cause des différences dans la composition chimique des structures qui réagissent différentiellement aux colorants; c'est la coloration.
Le comportement des structures tissulaires au cours de la coloration est fonction de leur composition chimique. Les colorants sont captés différemment par les différentes composantes tissulaires et apportent des contrastes sur une coupe. Par exemple, lorsqu'une composante quelconque absorbe beaucoup un colorant, sa densité optique augmente, réduisant l'amplitude de l'onde lumineuse passant au travers (lorsqu'observée au microscope); elle apparaît plus sombre que d'autres composantes absorbant moins le même colorant. De plus, en microscopie photonique, le colorant donne sa couleur aux composantes qui l'absorbent.
Il est maintenant coutumier d'utiliser deux colorants ou plus: un qui colore certaines composantes cellulaires, un autre qui colore d'autres composantes d'une couleur contrastante et, parfois, encore d'autres qui colorent d'autres composantes de couleurs différentes. Une combinaison souvent utilisée est l'hématoxyline-éosine (H&E).
L'hématoxyline oxydée (hématéine) et liée à un mordant (cation métallique: aluminium, fer, chrome, tungstène ou autre) est un colorant basique qui colore en bleu pourpre les composantes cellulaires basophiles (ayant une affinité pour les substances basiques), donc les composantes acides. Dans le noyau, le nucléole (contenant DNA et RNA) et la chromatine (d'apparence granulaire dans le nucléoplasme et contenant du DNA) sont colorés bleu pourpre. La membrane nucléaire apparaît colorée, mais c'est la chromatine adhérant à sa surface interne qui capte le colorant, non la membrane même. Dans le cytoplasme, les ribosomes, riches en RNA, sont colorés bleu pourpre; ces granules sont particulièrement mis en évidence dans les cellules ayant un taux élevé de synthèse protéique.
L'éosine liée à un radical anionique est un colorant acide qui imprègne une couleur rose rouge aux composantes cellulaires acidophiles (ayant une affinité pour les acides), donc les composantes basiques. Ce sont surtout les protéines structurales qui captent l'éosine et leur degré de coloration est affecté par le pH de la solution de colorant (les protéines -amphotères- se comportent en acide ou en base selon les conditions).
Les méthodes de coloration ne sont pas toutes basées sur la réaction du colorant avec des molécules de charges opposées. Des réactions chimiques connues peuvent être utilisées pour étudier les composantes tissulaires; c'est la science de l'histochimie. Par exemple, la coloration du glycogène par la technique de l'acide periodique-Schiff (APS ou PAS).
La plupart des colorants sont solubles dans l'eau, parfois dans l'alcool, mais non dans la paraffine. Les coupes montées sur lames et séchées doivent donc, préalablement à la coloration, être déparaffinées dans le xylène ou autre, réhydratées dans des bains d'éthanol successivement moins concentré, puis d'eau. Elles peuvent maintenant être colorées en une ou plusieurs étapes, selon la coloration choisie. La coloration est choisie en fonction du tissu et, comme il est à peu près impossible de pouvoir tout dévoiler à la fois, de la composante particulière du tissu qui nous intéresse dans le cadre de l'étude effectuée.
1.1.2.5 Montage permanent (entre lame et lamelle)
Pour conserver les coupes colorées, il faut les recouvrir d'une mince lamelle de verre ou de plastique scellée à l'aide d'un genre de colle d'indice réfractaire approprié. Ce milieu de montage n'étant pas soluble dans l'eau, il faut, immédiatement suite à la coloration, à nouveau déshydrater les coupes et les passer au xylène avant d'appliquer la lamelle.
Outre les colorations dont les principes viennent d'être exposés, il existe d'autres procédés de coloration utilisés en histologie. La coloration intravitale (aussi appelée coloration vitale) consiste à injecter un colorant à un animal vivant qui sera sacrifié plus tard. Pendant le temps de survie, le colorant se fixe sur certaines structures, par exemple le vert de Janus qui colore les mitochondries. Le colorant peut être phagocyté par des cellules qui le reconnaissent comme matière étrangère, par exemple le bleu Trypan ou l'encre de Chine retrouvés dans les macrophages. Il ne s'agit pas de coloration véritable, puisqu'il n'y a pas de liaison chimique entre les constituants cellulaires et le colorant, mais une rétention de celui-ci dans certaines cellules. La coloration supravitale s'applique à des cellules vivantes retirées de l'organisme, comme un frottis sanguin. Les réactions histochimiques, déjà mentionnées, permettent de localiser une substance chimique dans le tissu. L'imprégnation métallique consiste à laisser se déposer sur certaines structures tissulaires des atomes de métaux lourds (argent, chrome, or) qui les rendent opaques. Le métal recouvre la surface de la structure, il ne la pénètre pas.
1.2 DIFFERENCIATION CELLULAIRE ET DEVELOPPEMENT DES QUATRE TISSUS DE BASE
Une cellule unique, l'oeuf fertilisé (zygote), est l'ancêtre de toutes les cellules du corps. De ces millions de cellules, on reconnaît une centaine de types différents au microscope. Ainsi, deux processus importants se produisent au cours du développement: une prolifération cellulaire intense, requérant des millions de mitoses, et une différenciation cellulaire, qui fait que certaines cellules deviennent différentes d'autres cellules.
En embryologie, la notion de potentialité se réfère aux capacités non encore réalisées d'une cellule, ses possibilités. Elle dénote jusqu'à quel point une cellule peut servir d'ancêtre à différentes sortes de cellules (et non pas nombre de cellules). Puisque l'oeuf fécondé est l'ancêtre de toutes les sortes de cellules de l'organisme, il est totipotent. Au fur et à mesure que se déroulent les divisions mitotiques, des cellules-filles deviennent graduellement spécifiées ou déterminées. En acquérant une destinée différente de celle de la cellule-mère, elles diminuent leur potentialité; c'est le processus de différenciation. Au stade embryonnaire appelé gastrula, les trois feuillets tissulaires fondamentaux des métazoaires sont établis: ectoderme, mésoderme et endoderme.
La différenciation (détermination, spécification) des cellules de chaque feuillet se poursuit; elle n'affecte pas toutes les lignées cellulaires en même temps: chez un embryon d'âge donné, on retrouve des cellules à divers stades de différenciation. Elle tient du fait que certains gènes soient exprimés et d'autres supprimés, processus souvent réglé par le phénomène d'induction. Seules les cellules germinales, qui deviendront oeuf et spermatozoïde, (en contraste avec les cellules somatiques diploïdes) retiennent leur totipotentialité. Toutes les cellules somatiques n'atteignent pas le même degré de spécialisation. Certaines se spécialisent tellement qu'elles perdent la capacité de se diviser. Ce sont donc des cellules post-mitotiques.
Voici les quatre tissus de base chez l'adulte: épithélial
conjonctif
musculaire
nerveux
Epithélium désigne feuillet qui recouvre. Il comprend tous les feuillets de l'organisme qui recouvrent ou tapissent et qui sont constitués de cellules contiguës. La partie épithéliale de la peau dérive de l'ectoderme embryonnaire. Les épithélia tapissant les voies respiratoires et gastro-intestinales dérivent de l'endoderme embryonnaire. Celui tapissant la cavité péritonéale dérive du mésoderme embryonnaire. L'épithélium tapissant les vaisseaux sanguins et le coeur dérive aussi du mésoderme. Donc, le tissu épithélial adulte a des origines embryonnaires multiples.
Le tissu conjonctif dérive du mésoderme et comprend 5 classes:
tissu conjonctif ordinaire
tissu adipeux
cartilage
os
sang et tissu myéloïde.
Le tissu musculaire dérive aussi du mésoderme et comprend 3 classes:
lisse
cardiaque
strié squelettique
Le tissu nerveux, bien qu'une classe de tissu en soi, est essentiellement un épithélium modifié et spécialisé qui dérive d'une portion spécifiée de l'ectoderme embryonnaire: le neurectoderme. Il comprend le système nerveux central et le système nerveux périphérique.
Presque tous les organes se composent de plusieurs tissus.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}