CHAPITRE 10: SYSTEME CIRCULATOIRE
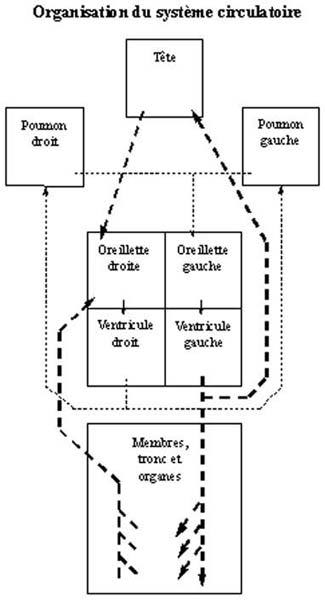
Voir schéma sur l'organisation du système circulatoire
Dans un organisme complexe comprenant plusieurs organes effectuant des fonctions spécialisées mais qui doivent être coordonnées pour assurer un ensemble harmonieux, un système de transport de matériels d'un organe à l'autre s'impose. Ce système de transport est le système circulatoire, et le sang qui y circule sert de véhicule à ce qui doit être transporté.
Mouvoir le sang dans les conduits requiert une pompe. Le coeur accomplit la fonction de deux pompes en parallèle, du moins chez les vertébrés supérieurs. Chaque pompe est reliée à un circuit. D'une part, la pompe droite pompe le sang désoxygéné dans le circuit pulmonaire et maintient la circulation pulmonaire. D'autre part, la pompe gauche pompe le sang oxygéné dans le reste de l'organisme et maintient la circulation systémique. Chez les vertébrés inférieurs, une pompe unique pourvoit à un système circulatoire plus simple.
Chaque pompe se divise en deux compartiments: l'oreillette où est collecté le sang qui revient au coeur et le ventricule qui reçoit le sang de l'oreillette homolatérale et l'envoie dans l'un des circuits. Le sang fraîchement oxygéné des poumons se jette dans l'oreillette gauche et le sang vicié en provenance des autres organes se jette dans l'oreillette droite.
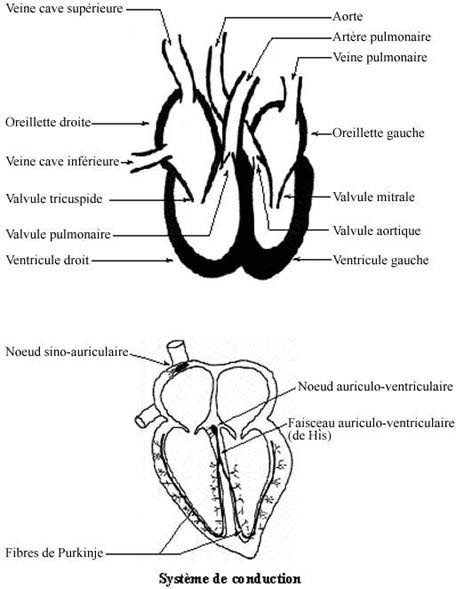
Des valves séparent les oreillettes des ventricules: la valvule tricuspide sépare l'oreillette droite du ventricule droit et la valvule mitrale sépare l'oreillette gauche du ventricule gauche. Entre deux battements cardiaques, ces valvules s'ouvrent pour permettre aux oreillettes de déverser leur contenu dans les ventricules respectifs. Elles se referment une fois le sang dans les ventricules et que ceux-ci se contractent. Les contractions ventriculaires font ouvrir deux autres valvules: la valvule pulmonaire, séparant ventricule droit et artère pulmonaire, et la valvule aortique, séparant ventricule gauche et tronc aortique. La haute pression infligée à ces grosses artères quand le sang s'y décharge fait refermer les deux valvules, ce qui empêche le retour du sang dans les ventricules. La pression est nettement plus élevée dans la circulation systémique que dans la circulation pulmonaire. Les grosses artères envoient le sang dans les embranchements de moindre calibre, les artères moyennes, qui distribuent le sang aux organes.
Les artères moyennes sont reliées à de nombreux embranchements de diamètre progressivement plus réduit mais à paroi puissante, les artérioles. Celles-ci opèrent comme une valve qui réduit la pression du sang qui les traverse avant d'atteindre des conduits à paroi mince, les capillaires. Les capillaires pulmonaires larguent dans l'air inspiré le CO2 sanguin et en soutirent l'O2 (revoir 6.1.1). Les capillaires de la circulation systémique livrent l'O2 aux tissus et leur en extirpent le CO2. Le sang des capillaires de chaque circuit poursuit sa course dans des vaisseaux de plus gros calibre, les veinules, qui se rejoignent pour former des veines de plus en plus grosses. Les veines ramenant le sang oxygéné de la circulation pulmonaire, les veines pulmonaires, se déversent dans l'oreillette gauche, tandis que les veines ramenant le sang désoxygéné de la circulation systémique, les veines caves, sont tributaires de l'oreillette droite.
La paroi du coeur et de la plupart des vaisseaux comprend trois couches dont l'épaisseur relative et la constitution varient avec la fonction particulière accomplie par la structure, ce qui permet de décrire un plan d'organisation histologique du système circulatoire:
|
localisation
|
coeur
|
vaisseaux
|
composition
|
|---|---|---|---|
|
interne
|
endocarde
|
tunica
intime
|
épithéliale
et conjonctive
|
|
moyenne
|
myocarde
|
tunica
media
|
musculaire
et conjonctive
|
|
externe
|
épicarde
|
tunica
adventitia
|
surtout
conjonctive
|
10.1 COEUR
La paroi des oreillettes et des ventricules comprend les trois couches nommées ci-haut. La paroi des oreillettes est plus mince que celle des ventricules correspondants et les parois de l'oreillette et du ventricule gauches sont plus épaisses que celles de leurs contreparties droites.
10.1.1 ENDOCARDE
L'endocarde tapisse les quatre compartiments cardiaques. Son épaisseur est en général inversement proportionnelle à celle du myocarde. De l'intérieur vers l'extérieur, il comprend:
- un endothélium d'une assise unique de cellules pavimenteuses ou polyhédrales
- une mince couche de tissu conjonctif collagénique et élastique avec ses fibroblastes
- une couche de tissu fibro-élastique à forte teneur en élastine et d'un peu de muscle lisse
- une couche conjonctive collagénique et adipeuse renfermant des vaisseaux sanguins et nerfs, équivalente à l'épimysium des muscles striés squelettiques (revoir 7.1).
Les deux premières couches de l'endocarde sont continues avec la tunica intima des gros vaisseaux reliés au coeur. Les valvules consistent en replis de l'endocarde renforcés de tissu conjonctif dense particulièrement élastique.
10.1.2 MYOCARDE
La plus grosse partie du coeur, particulièrement épaisse dans le ventricule gauche, le myocarde se compose surtout de tissu musculaire cardiaque (revoir 7.2). Le muscle cardiaque est séparé de l'endocarde d'une part et de l'épicarde d'autre part par du tissu conjonctif équivalent à l'épimysium du muscle squelettique. Continu avec celui-ci, un équivalent de périmysium divise le myocarde en faisceaux et sert de couloir aux vaisseaux sanguins irriguant le coeur (vaisseaux coronariens) et aux nerfs l'innervant. Finalement, un équivalent d'endomysium composé de fines fibres collagéniques, élastiques et réticulaires dans la substance amorphe, entoure chaque fibre cardiaque et lui apporte capillaires sanguins, lymphatiques et fibres nerveuses. Somme toute, le tissu conjonctif est relativement peu abondant dans le coeur en bonne santé.
L'efficacité du coeur en tant que pompe double dépend de la simultanéité des contractions des côtés gauche et droit ainsi que de la synchronisation des séquences du cycle cardiaque. Cette efficacité incombe à un système d'amorce et de conduction spécialisé pour la fonction cardiaque, assuré par un type cellulaire du myocarde différent des cardiomyocytes typiques mais qui dérive de leur spécialisation. Plus petites que les cardiomyocytes, les cellules de ce système sont spécialisées pour l'amorce et la propagation d'une vague de dépolarisation, plutôt que pour la contraction. Elles renferment peu de myofibrilles mais beaucoup de glycogène. Elles s'agrègent en petites masses enrobées de tissu conjonctif fibroélastique pourvu de nombreux capillaires. Ce système de conduction comprend:
le noeud sino-auriculaire: dans la paroi de la veine cave supérieure à sa jonction avec l'oreillette droite; il constitue le pacemaker du coeur.
le noeud auriculo-ventriculaire: à la jonction des oreillettes et ventricules, et qui se poursuit en faisceau auriculo-ventriculaire.
le faisceau auriculo-ventriculaire (de His): dans la paroi interventriculaire, continu avec les fibres de Purkinje.
les fibres de Purkinje: autour de chaque ventricule.
La vague de dépolarisation est amorcée dans le noeud sino-auriculaire. Les deux divisions du système nerveux viscéral (orthosympathique et parasympathique) modifient mais ne déclenchent pas la dépolarisation; celle-ci est une propriété intrinsèque du noeud. Du noeud sino-auriculaire, la vague de dépolarisation se transmet à tous les cardiomyocytes des oreillettes puis converge vers le noeud auriculo-ventriculaire. De ce noeud, la vague passe au faisceau de His et aux fibres de Purkinje, qui la transmettent simultanément à toutes les cellules musculaires des ventricules. Ainsi, ce système de conduction rapide permet la contraction simultanée de toutes les parties des oreillettes lorsqu'elles déversent leur sang dans les ventricules, puis des ventricules lorsqu'à leur tour ils déversent leur sang dans l'aorte et l'artère pulmonaire.
10.1.3 EPICARDE
Couche de tissu conjonctif fibroélastique recouverte d'un mésothélium: feuillet épithélial de cellules pavimenteuses ou polyhédrales.
L'épicarde est lui-même recouvert d'un autre feuillet mésothélial appelé péricarde, duquel il est séparé par la cavité péricardiaque remplie d'un film de fluide. Epicarde et péricarde sont continus à la racine des grands vaisseaux. Le péricarde n'est pas considéré comme appartenant au coeur mais aux mésentères enveloppant les organes des cavités thoracique et abdominale.
10.2 ARTERES ET ARTERIOLES
Le sang provenant du coeur est conduit aux différents organes par les artères et leurs embranchements. A chaque niveau du système circulatoire, les artères jouent un rôle quelque peu différent. Leur rôle respectif se traduit par l'épaisseur relative de leur paroi et leur composition histologique. Dans l'ensemble, la tunica media est la couche la plus importante des artères.
10.2.1 ARTERES ELASTIQUES (Ex: aorte, artère pulmonaire, artère rénale)
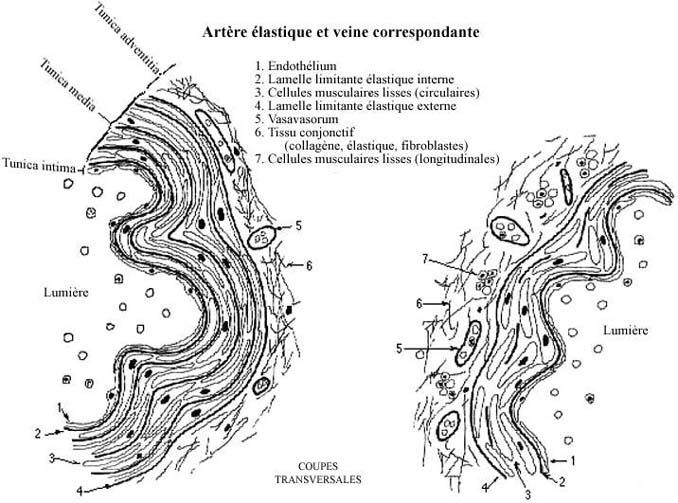
Voir schémas de l'artère élastique et la veine correspondante
Les ventricules livrent le sang aux grandes artères sous forte pression (pression systolique). Pour accomoder une telle pression, la paroi de ces artères doit pouvoir être étirée. La présence d'une quantité importante d'élastine dans la paroi permet pareil étirement. De plus, en retournant à leur longueur de repos, les fibres élastiques se trouvent à exercer une pression sur le flot sanguin, le poussant plus distalement dans les artères en le maintenant sous pression. Les trois couches des artères élastiques dénotent cette fonction.
La tunica intima comprend un endothélium, une couche de cellules pavimenteuses à polyhédrales, et une lamelle limitante élastique interne, d'élastine et assez épaisse pour être visible au H&E. Même si la majorité des auteurs considèrent cette lamelle comme appartenant à l'intima, il serait plus logique de l'inclure dans la tunica media. L'endothélium des vaisseaux sanguins n'accomplit pas qu'une fonction structurale; en effet, les cellules endothéliales sécrètent un produit qui facilite la circulation du sang dans les vaisseaux.
La tunica media se compose de cellules musculaires lisses (revoir 7.3) disposées en feuillets circulaires, ou plutôt spiralés, en alternance avec des couches d'élastine qu'elles fabriquent elles-mêmes. (Rappelez-vous que le fibroblaste et la cellule musculaire lisse dérivent tous les deux de la cellule mésenchymale. Il n'est pas étonnant que la cellule musculaire lisse démontre à l'occasion des propriétés normalement rencontrées chez le fibroblaste, d'autant plus qu'elle est relativement moins spécialisée que, par exemple, la fibre musculaire striée squelettique.) La tunica media ne contient donc pas de fibroblastes. La media est délimitée par une couche d'élastine épaisse et visible appelée lamelle limitante élastique externe.
La tunica adventitia se compose de tissu conjonctif composé de fibres collagéniques et élastiques renfermant des petits vaisseaux sanguins, lymphatiques et nerfs desservant l'artère. On donne le nom de vasavasorum à un vaisseau sanguin localisé dans la paroi d'un autre vaisseau sanguin. Les cellules des couches les plus internes reçoivent O2 et substances nutritives à partir du sang circulant dans la lumière artérielle, par diffusion. Celle-ci ne pouvant s'effectuer que sur une distance limitée, les cellules en position plus externe requièrent donc un apport sanguin additionnel, assuré par les vasavasora de l'adventitia.
10.2.2 ARTERES MUSCULAIRES OU DE DISTRIBUTION (Ex: artères splénique, coronaires).
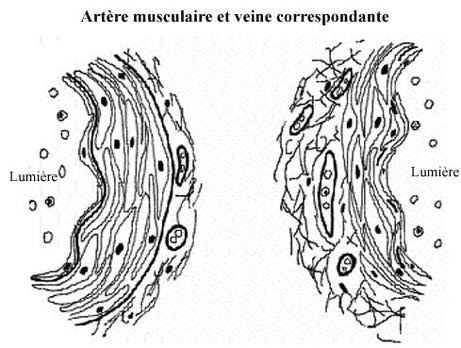
Voir schémas de l'artère musculaire et la veine correspondante
Les différents organes et parties de l'organisme n'éprouvent pas tous les mêmes besoins en sang. Par exemple, une petite glande comme la thyroïde ne demande pas le même volume sanguin qu'une glande aussi considérable que le foie. De plus, le besoin de chacun d'eux varie selon son niveau d'activité. Il va de soi que les muscles des jambes nécessitent un apport sanguin beaucoup plus considérable durant la course que pendant le sommeil. Il en va de même pour l'estomac en pleine activité de digestion. Une façon d'ajuster l'apport sanguin aux besoins individuels et ponctuels des organes est de modifier le calibre des artères qui distribuent le sang à ces organes et, de ce fait, régler la quantité de sang qui les atteint.
L'ajustement du diamètre artériel s'effectue par contraction musculaire et relève du système nerveux viscéral impliqué dans un jeu de rétroactions. La présence de nombreuses cellules musculaires lisses dans la paroi des artères de distribution, aussi appelées artères musculaires, permet pareille fonction. Le système nerveux peut les affecter, alors qu'il ne peut agir sur du matériel intercellulaire comme l'élastine des artères élastiques.
La tunica intima comprend un endothélium et une lamelle limitante élastique interne.
La tunica media, elle, se compose de cellules musculaires lisses et de beaucoup moins d'élastine que dans l'artère élastique. Un équivalent d'endomysium, très réduit, dirige les capillaires sanguins et les fibres nerveuses vers les myocytes. Une lamelle limitante élastique externe, généralement visible sans coloration spéciale, délimite la tunique moyenne.
La tunica adventitia, typique, abrite du tissu conjonctif collagénique et élastique, des vasavasora et des faisceaux de nerfs périphériques.
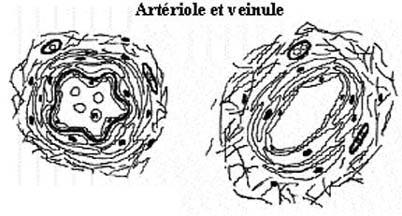
10.2.3 ARTERIOLES
Voir schémas de l'artériole et de la veinule
Une fois le sang artériel distribué aux organes par les artères musculaires, les artérioles en réduisent la pression pour le livrer à des vaisseaux à mince paroi sans briser celle-ci. Les artérioles se caractérisent par une lumière relativement faible et une paroi relativement épaisse; leur diamètre total ne dépasse généralement pas 100µm, dont la lumière compte pour environ la moitié. Le ratio lumière/paroi était souvent plus élevé dans les artères. Les trois tuniques se retrouvent dans la paroi artériolaire.
La tunica intima consiste en un endothélium et une lamelle limitante élastique interne encore bien visible. Celle-ci est typiquement dentelée, faisant ressortir les noyaux des cellules endothéliales dans la lumière.
La tunica media comprend quelques couches de cellules musculaires lisses spiralées avec peu ou pas d'élastine. Le système nerveux viscéral contrôle leur degré de contraction. Les hormones influencent aussi leur tonus. Une élévation anormale du tonus musculaire des artérioles résulte en une condition d'hypertension. La lamelle limitante élastique externe est généralement absente.
La tunica adventitia, mince puisque les artérioles empruntent déjà le parcours formé par le tissu conjonctif de l'organe récipiendaire, consiste surtout en collagène et peut ne pas renfermer de vasavasora.
10.3 CAPILLAIRES
Sites des échanges des gaz respiratoires, substances nutritives et déchets métaboliques entre le sang qu'ils renferment et le compartiment extrasanguin du tissu qu'ils parcourent, les capillaires réduisent l'épaisseur de leur paroi au strict minimum. Un mince endothélium de cellules pavimenteuses et de délicates fibres conjonctives fournies par le tissu récipiendaire rendent les échanges efficaces. Le diamètre moyen d'un capillaire est de 8 à 10µm. Dans certains organes, notamment les organes lymphatiques et la moelle osseuse, des capillaires à endothélium particulièrement mince et lumière agrandie prennent le nom de sinusoïdes. Dans les capillaires un peu plus gros, de petites cellules musculaires lisses peuvent entourer l'endothélium. On retrouve souvent des péricytes (revoir 3.2.2) à la surface des capillaires; ils ne répondent pas à l'influx nerveux et ne se contractent pas.
Quant à la morphologie des cellules endothéliales et à la fonction qu'elle sous-tend, on décrit deux types de capillaires: continus et discontinus. Quant à la complexité de leur ramification, l'on peut aussi décrire quelques types capillaires, aux deux extrêmes se trouvant les réseaux capillaires et les anastomoses artério-veineuses.
10.3.1 CAPILLAIRES CONTINUS versus CAPILLAIRES DISCONTINUS OU À HUBLOTS
Voir schémas de ces capillaires.
Dans les capillaires continus, la membrane de cellules endothéliales adjacentes décrit des interdigitations, un peu comme dans les disques intercalaires, sites de jonctions cellulaires serrées de type occludens. Comme pour tout épithélium, une membrane basale souligne l'endothélium capillaire. Un tel arrangement ne permet pas un échange facile des substances entre les compartiments sanguin et extrasanguin; la pinocytose et l'exocytose sont nécessaires pour permettre aux substances autres que gazeuses de traverser l'endothélium. Les capillaires continus conviennent aux organes particulièrement exposés aux microbes extérieurs, tel le poumon, ou aux organes où il est extrêmement critique que le passage de substances soit très sélectif et contrôlé, tel le système nerveux central.
Dans le cas des capillaires discontinus, les jonctions entre les cellules endothéliales sont moins serrées, de type adherens, et le cytoplasme est perforé de petites fenêtres, ou hublots, permettant le passage rapide et moins sélectif de certaines substances. Dans la plupart des capillaires discontinus, des diaphragmes, dont la nature demeure énigmatique à nos jours, obstruent partiellement les hublots. Toutefois, dans le rein, où le transport s'effectue à un rythme excessif, les hublots ne sont pas refermés par des diaphragmes et la membrane basale est particulièrement mince ou absente.
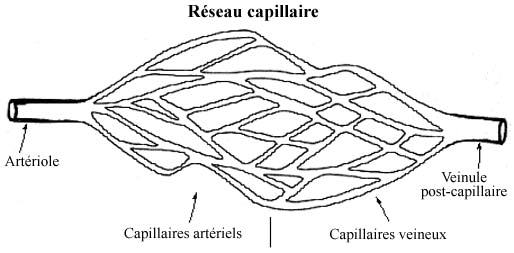
10.3.2 RESEAUX CAPILLAIRES versus ANASTOMOSES ARTERIO-VEINEUSES
Dans la plupart des organes, les capillaires artériels forment un réseau plus ou moins élaboré d'embranchements anastomosés. Dans tel réseau les capillaires artériels deviennent éventuellement capillaires veineux. C'est au niveau du réseau que s'opèrent les échanges des gaz respiratoires et substances diverses entre le sang et les tissus, que le métabolisme de la substance amorphe est équilibré entre le compartiment sanguin et l'espace intercellulaire, et que se forme la lymphe par l'excédent de plasma exsudé des capillaires veineux (revoir 3.1.2).
Une anastomose artério-veineuse se dit d'un capillaire qui passe directement d'artériel à veineux sans de nombreux embranchements intermédiaires. Les capillaires des anastomoses tendent à avoir un diamètre légèrement plus grand que les capillaires des réseaux. Les anastomoses se rencontrent dans les organes où les capillaires ne jouent pas qu'un rôle de substrat pour échanges de substances mais également un rôle dans la thermorégulation. Les capillaires de la peau et des voies respiratoires offrent des exemples de choix.
10.4 VEINULES ET VEINES
Le sang ayant circulé dans le système artériel, maintenant modifié selon qu'il appartient au circuit pulmonaire ou systémique, retourne au coeur par le système veineux en empruntant, la plupart du temps, un chemin parallèle mais en sens inverse. A chaque vaisseau du système artériel correspond un vaisseau du système veineux: capillaires veineux, veinules, veines moyennes et grosses veines.
Les veinules et veines se distinguent respectivement des artérioles et artères par un taux lumière/paroi beaucoup plus élevé et par l'ampleur relative de la tunica adventitia, au détriment de la média. De plus, l'élastine prend moins d'importance, supplantée par du collagène, et il est rare de reconnaître une membrane limitante élastique externe. A cause de la faible teneur en O2 du sang qui circule dans les veinules et veines systémiques, elles renferment plus de vasavasora dans leur adventitia que les artérioles et artères correspondantes ou que les veinules et veines du circuit pulmonaire.
Bien qu'effectivement moins musculaires, certaines veines, notablement les grosses veines travaillant contre la gravité (grosses veines des pattes, veine cave inférieure), renferment une bonne quantité de muscle, notamment des faisceaux de muscles lisses longitudinaux dans leur tunica adventitia, et même des valves qui empêchent la circulation à rebours. Ces valves consistent en replis de la tunica intima renforcés de tissu conjonctif élastique.
10.5 CIRCULATION LYMPHATIQUE
Des capillaires incorporant la lymphe, plutôt que du sang, se forme in situ dans presque tous les organes. Les capillaires lymphatiques ne sont pas en continuité anatomique avec les capillaires sanguins (artériels ou veineux) mais constituent un circuit parallèle. Les capillaires lymphatiques se regroupent pour former des vaisseaux lymphatiques de plus gros calibre qui quittent les organes et, en rejoignant d'autres vaisseaux lymphatiques, forment des vaisseaux de plus en plus gros. La lymphe est finalement drainée dans deux troncs lymphatiques, le canal thoracique et le canal lymphatique droit (il n'existe pas de canal lymphatique gauche, du moins chez l'humain) qui se déversent dans la circulation veineuse.
Les capillaires lymphatiques possèdent une paroi distinctive en ce que leur endothélium n'est accompagné d'aucune membrane basale ni de péricytes. Leur diamètre dépasse généralement celui des capillaires sanguins. Les vaisseaux lymphatiques plus gros comprennent les trois tuniques, similaires à celles des veines et peu musculaires, et leur paroi est munie de valves.
Aucune pompe (analogue au coeur) n'inflige de pression à la lymphe. La circulation lymphatique est contrôlée par l'action d'hormones et du système nerveux viscéral sur les myocytes (peu abondants) de leur paroi et est facilitée par l'activité générale de l'organisme. La présence de valves dans la paroi des vaisseaux lymphatiques empêche la circulation à rebours.
La lymphe se compose de fluide analogue au plasma sanguin, mais dont la composition biochimique diffère, notamment par sa teneur en lipides qui excède celle du sang. La composition de la lymphe varie également d'une région à l'autre de l'organisme, soit selon son site d'origine. Elle s'enrichit de l'excès de fluide extracellulaire, capté par les capillaires lymphatiques. Elle contient principalement un type cellulaire, les lymphocytes, mais aussi une certaine quantité de monocytes.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}