CHAPITRE 17: SYSTEME REPRODUCTEUR FEMELLE
Le système reproducteur femelle comprend deux ovaires, deux oviductes, un utérus, un vagin (utérus et vagin sont pairs ou bicornes chez les marsupiaux), les parties génitales externes et les glandes mammaires, chez les mammifères, de nombre variable avec les espèces.
17.1 OVAIRE
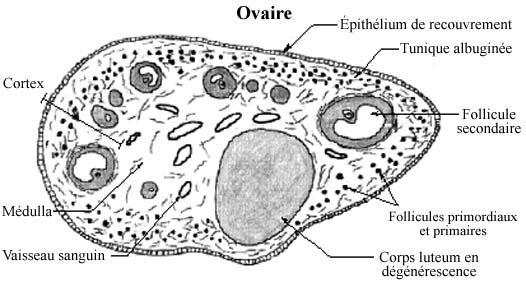
Localisé dans la paroi latérale de la cavité pelvienne, l'ovaire a la forme d'une amande. Au lieu d'être enveloppé d'une capsule conjonctive comme les autres organes, il est recouvert d'un épithélium simple cuboïdal ou cylindrique. Une capsule conjonctive, même mince, ne permettrait pas facilement l'ovulation (voir plus bas).
En coupe, on distingue deux régions principales: le cortex, plus important, et la médulla au centre. Juste sous l'épithélium de recouvrement, une zone du cortex contient peu de cellules et beaucoup de matériel intercellulaire lâche, c'est la tunique albuginée de l'ovaire. Sous elle, la densité cellulaire augmente, comme on s'y attend du cortex d'un organe. Cette région renferme les follicules ovariens. La médulla comprend du tissu conjonctif lâche, dont une bonne proportion de fibres élastiques, des cellules musculaires lisses, des artères (spiralées) et veines en route vers le cortex.
Les cellules de la lignée germinale (gonocytes) femelles, les ovogonies, se divisent mitotiquement pour atteindre un nombre très élevé durant la vie foetale, mais qui régresse postnatalement. En accroissant sa taille, l'ovogonie devient ovocyte primaire, d'environ 20µm de diamètre. Chacun s'entoure d'un épithélium simple pavimenteux composé de cellules dites folliculaires; ovocyte I et épithélium folliculaire simple pavimenteux forment ensemble le follicule primordial. Le follicule entier mesure environ 35µm de diamètre; les cellules folliculaires sont donc beaucoup plus petites que l'ovocyte. L'ovocyte entreprend une première division méiotique qui s'arrête au stade prophase et y demeure jusqu'à la puberté. Son cytoplasme abondant et pâle contient des granules vitellins.
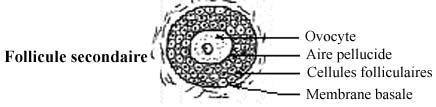
A la puberté et ensuite, les hormones hypophysaires gonadotrophes FSH et LH (voir 18.1.1.1) stimulent le développement d'un certain nombre de follicules selon un cycle précis, nombre de follicules et durée du cycle variant selon les espèces animales. Le développement folliculaire se traduit par l'augmentation de la taille des cellules folliculaires qui, de pavimenteuses, deviennent cuboïdales puis cylindriques, et par leur multiplication qui résulte en la formation d'un épithélium stratifié. L'ovocyte augmente sa taille et produit une épaisse membrane basale appelée zona pellucida, à la formation de laquelle les cellules folliculaires immédiatement autour de l'ovocyte contribuent peut-être. La membrane cytoplasmique de l'ovocyte et celle des cellules folliculaires décrivent des microvillosités qui pénètrent la zona pellucida. Le follicule dont l'épithélium est simple cuboïdal ou cylindrique est appelé follicule primaire et celui dont l'épithélium est stratifié follicule secondaire. La petite taille des cellules folliculaires et leur densité donnent une apparence granulaire à l'épithélium folliculaire stratifié, qui se voit appelé zona granulosa.
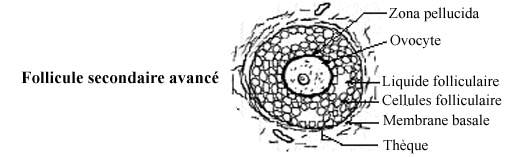
Puisque le tissu épithélial est invasculaire, il n'y a pas de capillaires sanguins parmi les cellules folliculaires; ils se retrouvent dans le conjonctif entre les follicules. Les cellules folliculaires sont donc nourries et oxygénées par diffusion à partir de ces capillaires, et l'ovocyte par diffusion au travers l'épithélium folliculaire, qui agit comme barrière protectrice. Autour des follicules, des cellules et fibres conjonctives se disposent de façon concentrique, formant une thèque (enveloppe), encore peu développée. Une membrane basale sans nom spécial, et moins épaisse que la zona pellucida, sépare l'épithélium folliculaire de la thèque.
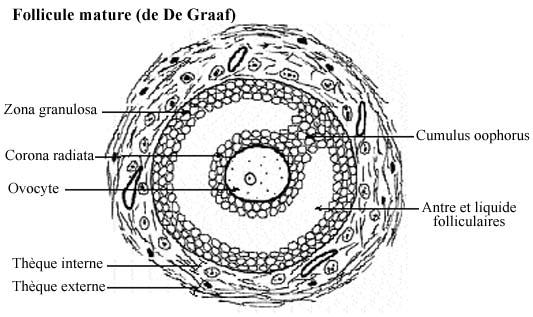
Les cellules folliculaires se multiplient et sécrètent un fluide folliculaire qui s'accumule dans l'espace entre elles, formant comme des petits lacs (voir follicule secondaire avancé). Du plasma exsudé des capillaires interfolliculaires s'y ajoute. Graduellement, les lacs de fluide folliculaire fusionnent en un grand bassin, l'antre folliculaire, qui divise l'épithélium folliculaire en deux régions: quelques couches cellulaires restent autour de l'ovocyte, la corona radiata, les autres couches entourant l'antre; ces deux régions demeurent reliées par un pont de cellules folliculaires: le cumulus oophorus. Donc, l'antre folliculaire n'entoure pas complètement la corona radiata. Certains auteurs incluent la corona radiata dans le terme cumulus oophorus.
Dans le follicule mûr, ou de De Graaf, la thèque se divise en deux régions. La thèque interne, pourvue d'abondants capillaires, contient des cellules conjonctives modifiées, ovoïdes ou arrondies, dont le cytoplasme renferme des gouttelettes lipidiques. La thèque externe, moins vascularisée, se compose de fibroblastes, fibres conjonctives et cellules musculaires lisses.
Dans l'ovaire d'une femelle sexuellement mûre, il se retrouve des follicules à tous les stades de développement. Les follicules primordiaux et primaires tendent à occuper la périphérie du cortex, sous la tunique albuginée, et les follicules secondaires la région profonde du cortex. Les follicules mûrs sont si gros, 10 à 12mm de diamètre, qu'ils occupent toute l'épaisseur corticale et créent même une protubérance à la surface de l'ovaire. Plusieurs follicules n'atteignent pas la maturité et régressent (follicules atrésiques).
Une quantité considérable de fluide est produite dans le follicule mûr, provenant surtout du plasma exsudé des capillaires de la thèque et biochimiquement modifié par les sécrétions des cellules folliculaires et des cellules de la thèque. Le fluide exerce une grande pression sur l'épithélium folliculaire, les thèques et également sur la tunique albuginée et l'épithélium de recouvrement ovarien. Ces barrières cèdent éventuellement et le fluide s'écoule dans la cavité péritonéale; le cumulus oophorus se détache et l'ovocyte entouré de sa corona radiata, soit l'ovule, est livré dans la cavité péritonéale: c'est le phénomène d'ovulation ou ponte ovulaire. L'ovocyte complète alors la première division méiotique entamée vers la fin de la vie foetale. La suite de la maturation ovocytaire ne sera pas considérée dans ce cours.
Après l'ovulation, le reste du follicule dans l'ovaire se développe en corpus luteum, ou corps jaune: les cellules folliculaires restantes et de la thèque interne, appelées cellules interstitielles et analogues à celles du testicule, sous l'effet de la LH sécrètent des hormones oestrogènes, activité amorcée avant l'ovulation. Les cellules interstitielles adoptent une organisation en cordons, séparés par des capillaires sanguins, comme plusieurs glandes endocrines. Parmi les hormones sécrétées, la progestérone agit, via la circulation sanguine, sur la paroi utérine et la prépare pour l'implantation éventuelle de l'ovocyte fertilisé. Si l'implantation ne se produit pas, le corps jaune régresse graduellement, formant le corpus albicans, ou corps blanc. Ainsi privé de stimulation, l'utérus réagit par une manifestation appelée menstruations, d'expression variable chez les différentes espèces.
17.2 OVIDUCTE
L'extrémité de l'oviducte (trompe utérine ou de Fallope), agrandie en entonnoir, recouvre l'ovaire sans être continue avec lui. L'ovule expulsé de l'ovaire est ainsi capté par l'oviducte et conduit à l'utérus, dans lequel aboutissent les oviductes. La fécondation se produit dans l'oviducte (chez les animaux à fécondation interne). Chez des mammifères, l'implantation a lieu normalement dans l'oviducte, chez d'autres dans l'utérus.
La surface interne de l'oviducte, un peu comme celle de l'uretère, dessine plusieurs replis. La paroi comprend trois couches: muqueuse, musculaire et séreuse. L'épithélium simple cylindrique comprend des cellules ciliées et des cellules muqueuses, plus ou moins en alternance. Son épaisseur varie avec le cycle menstruel. La lamina propria, de tissu conjonctif lâche, réagit comme l'endomètre utérin (voir 17.3) en cas d'implantation. La musculaire, avec sa couche interne de fibres musculaires lisses circulaires et sa couche externe de fibres longitudinales (comme dans l'intestin), permet un certain mouvement péristaltique, accéléré au temps de l'ovulation et qui, avec le battement des cils, contribue à l'avancement de l'ovule. La paroi de l'oviducte peut contenir des lymphocytes et nodules lymphatiques.
17.3 UTERUS
Site de l'implantation chez plusieurs mammifères, l'utérus a une paroi épaisse qui se modifie avec le cycle hormonal et selon qu'il est gravide ou non. Trois couches s'y reconnaissent: muqueuse appelée endomètre, musculaire appelée myomètre, et séreuse.
Le myomètre utérin consiste en faisceaux de muscle lisse séparés par du tissu conjonctif grandement élastique. Ces faisceaux forment trois plans vaguement définis: interne de fibres obliques et externe de fibres longitudinales, les deux minces, puis moyen de fibres circulaires, épais et qui contient plusieurs vaisseaux sanguins (stratum vascularae). L'épaisseur du myomètre peut décupler durant la grossesse: non seulement les myocytes s'hypertrophient-ils, mais de nouveaux myocytes se forment à partir de ceux existants ou de péricytes (revoir 3.2.2).
L'endomètre change considérablement au cours du cycle hormonal. Son épithélium simple de cellules ciliées et muqueuses s'invagine dans les couches profondes pour former des glandes simples tubulaires muqueuses, dont le degré d'activité varie avec le cycle. Sa lamina propria épaisse et richement vascularisée est continue avec le myomètre. On décrit plutôt l'endomètre en couche fonctionnelle, comprenant l'épithélium et une bonne partie des glandes et de la lamina propria, et couche basale, portion profonde des glandes et de la lamina propria ancrée au myomètre. La couche fonctionnelle est désintégrée et rejetée lors des menstruations; elle participe à la formation du placenta quand se produit l'implantation. La couche basale est à l'origine de sa regénération.
La portion inférieure de l'utérus, étroite et continue avec le vagin, se distingue par sa composition pas tant musculaire que collagénique dense; il s'agit du col utérin (cervix). Son épithélium ressemble à celui du reste de l'utérus, excepté pour la région adjacente au vagin qui ressort dans celui-ci; cette région comprend un épithélium stratifié pavimenteux non kératinisé comme celui du vagin. Entre les deux types épithéliaux s'insère une courte zone de transition. A la parturition, le col utérin doit se dilater considérablement, dilatation qui s'opère par un amolissement de sa matière collagénique, présumément sous l'effet d'hormones peptidiques appelées relaxines.
17.4 VAGIN
Le vagin sert de passage à l'organe copulateur mâle pour l'accouplement. A la parturition, il sert de passage entre l'utérus, duquel il est gardé par le col, et le monde extérieur. Tube fibromusculaire ancré aux structures voisines par une adventitia, le vagin est dépourvu de glandes. L'épithélium qui le tapisse appartient au type stratifié pavimenteux ou cuboïdal non kératinisé, modifiable avec le cycle menstruel. La lamina propria de tissu conjonctif assez dense renferme des nodules lymphatiques. La musculaire comprend des fibres circulaires et longitudinales qui ne forment pas de plans distincts. Le vagin est abondamment pourvu de terminaisons nerveuses sensorielles.
17.5 PARTIES GÉNITALES EXTERNES
L'apparence des parties génitales externes varie beaucoup chez les différents animaux. Un patron très général peut être tracé pour les mammifères. L'entrée du vagin, le vestibule, est limitée par deux replis cutanés bourrés de tissu adipeux et de glandes muqueuses, les lèvres majeures, analogues au scrotum mâle. Elles se rejoignent antérieurement et, en cet angle, se situe une petite masse de tissu érectile, le clitoris, analogue au pénis mâle, mais non traversé par l'urètre. L'ouverture urétrale se trouve entre le clitoris et le vestibule. Le clitoris est recouvert d'un autre repli cutané, le prépuce, qui se poursuit postérieurement en lèvres mineures, parallèles et médianes aux lèvres majeures. Elles ont un recouvrement cutané intermédiaire entre celui du vagin et des lèvres majeures.
17.6 GLANDES MAMMAIRES (CHEZ LES MAMMIFERES)
Invaginations épithéliales d'une région de l'ectoderme embryonnaire appelée ligne de lait, les glandes mammaires consistent en glandes sudoripares modifiées. Chaque glande mammaire est composée alvéolaire, et sa sécrétion, le lait, peut se qualifier aussi bien de muqueuse que de séreuse. A la puberté, les glandes mammaires de la femelle grossissent, mais de façon plus ou moins marquée, selon les espèces. Ce grossissement s'accomplit par l'addition de tissu adipeux et conjonctif fibreux entre les canaux excréteurs. Les unités de sécrétion ne se développent qu'en réponse à un signal hormonal émis durant la gestation.
La glande mammaire au repos, i.e., d'une femelle sexuellement mûre mais ni gestante ni en lactation, comprend une tétine recouverte d'un épithélium stratifié pavimenteux kératinisé très mince et pigmenté. Au bout de la tétine émergent les canaux lactifères, pendants des canaux interlobaires des grosses glandes exocrines composées. Un épithélium comme celui de la tétine tapisse d'abord le canal lactifère, puis il s'amincit en deux couches de cellules cylindriques. Entre les canaux lactifères, formant la substance de la tétine, se trouvent du tissu conjonctif dense, des cellules musculaires lisses encerclant les canaux, des vaisseaux sanguins et des terminaisons sensorielles encapsulées.
Chaque glande mammaire comprend plusieurs lobes, délimités par des partitions conjonctives. Un lobe se définit comme étant le tissu glandulaire drainé par un canal lactifère. Chaque lobe comprend plusieurs lobules, délimités par du tissu conjonctif et drainés par les canaux intralobulaires. Avant d'aboutir dans les canaux lactifères, les canaux intralobulaires se dilatent en sinus lactifères qui servent de réservoir de la sécrétion. Sinus et canaux sont formés du même épithélium à deux assises cellulaires. Le tissu conjonctif qui remplit les espaces entre les canaux est la contrepartie de la couche papillaire du derme.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}