> Tissu conjonctif (3.1, 3.2, 3.3)
CHAPITRE 3: TISSU CONJONCTIF ORDINAIRE ET ADIPEUX
Le tissu conjonctif porte ce nom du fait qu'il soutient les autres tissus. Il tient ensemble les différentes parties du corps et donne une forme à celui-ci. Cette fonction de soutien est assurée par le matériel intercellulaire que produisent les cellules du tissu conjonctif, matériel pouvant être délicat ou très rigide. Ainsi, nous étudierons chaque classe de tissu conjonctif en décrivant ses éléments cellulaires, d'une part, et ses éléments extracellulaires, d'autre part. Plusieurs fonctions autres que le soutien sont aussi accomplies par certains types de tissu conjonctif. Nous avons déjà vu la classification du tissu conjonctif en 1.2:
- tissu conjonctif ordinaire
- tissu adipeux
- cartilage
- os
- sang et tissu myéloïde
Le tissu conjonctif ordinaire est grandement constitué de matériel intercellulaire élaboré par certaines cellules, surtout les fibroblastes. Ce matériel intercellulaire du tissu conjonctif ordinaire comprend deux composantes: fibreuse et amorphe.
3.1 MATERIEL INTERCELLULAIRE DU TISSU CONJONCTIF ORDINAIRE
3.1.1 COMPOSANTE FIBREUSE
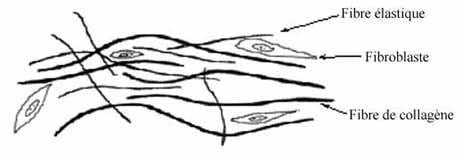
3.1.1.1 Fibres de collagène (grec: colla=colle; gennan=produire)
Les plus abondantes, les fibres collagéniques portent ce nom du fait qu'une fois bouillies, elles forment de la colle. Elles sont composées de la protéine collagène, une protéine très résistante. Ainsi, une pièce de viande contenant beaucoup de collagène est dure; bouillie, son collagène hydraté se convertit en gélatine. Elle devrait donc être bouillie pendant plusieurs heures pour devenir tendre plutôt qu'être rôtie. Le collagène peut être transformé en cuir: en digérant l'épithélium de la peau des animaux et en tannant le tissu conjonctif dense dermique et sous-cutané, grandement constitué de fibres collagéniques, ce tissu devient très résistant aux changements chimiques.
Dans le tissu conjonctif ordinaire, les fibres collagéniques sont produites par des cellules appelées fibroblastes. Le suffixe "blaste" suppose que la cellule a conservé le pouvoir de se diviser, tandis que "cyte" (fibrocyte) implique la fin d'une lignée cellulaire. Toutefois, les deux sont souvent employés sans discrimination. Les fibroblastes synthétisent des molécules de procollagène qu'ils sécrètent à leur surface, où elles sont converties en molécules de tropocollagène qui s'assemblent en fibrilles de collagène, lesquelles montrent une périodicité axiale de 64nm. Ces dernières s'apposent pour finalement former les fibres de collagène, qui peuvent atteindre jusqu'à 10µm de diamètre et qui ne sont pas embranchées. Les fibroblastes deviennent enrobés dans les fibres sécrétées. Les fibres de collagène se colorent en rose au H&E et peuvent être parfois confondues avec des fibres musculaires lisses. La coloration appelée trichrome de Masson les colore spécifiquement en vert turquoise.
3.1.1.2 Fibres d’élastine : propriété d'élasticité.
Les fibres
élastiques sont composées de la protéine élastine.
Dans le tissu conjonctif ordinaire elles sont sécrétées
par les fibroblastes. Ils synthétisent et sécrètent
des molécules de proélastine qui sont converties en tropoélastine
puis assemblées en desmosine et, finalement, en élastine. L'élastine
est entourée de microfibrilles composées de protéine microfibrillaire,
elle aussi synthétisée par les fibroblastes. Les fibres élastiques
ne comprennent pas de sous-unités fibrilles (d'élastine, par analogie
aux fibrilles de collagène), ni ne montrent de périodicité.
Elles sont embranchées et leur diamètre peut atteindre 1µm.
Les fibres élastiques ne se colorent pas bien au H&E mais leur qualité réfractaire les rend parfois visibles là où elles abondent. Elles se démontrent grâce aux colorations à l'orcéine ou à la résorcine-fuchsine, qui utilisent l'argent.
Les fibres collagéniques et élastiques peuvent être très longues et forment souvent des couches. Dans les tendons, les fibres collagéniques sont droites et parallèles, assurant la rigidité. Dans le derme de la peau, les fibres collagéniques sont sinueuses et les fibres élastiques, plus courtes, droites. La flexibilité de la peau, nécessaire pour tout mouvement, est attribuable aux fibres de collagène sinueuses qui, lors d'un mouvement d'étirement, deviennent droites sans être étirées (elles ne sont pas élastiques); ce sont elles qui déterminent la limite d'étirement du tissu. Les fibres élastiques, droites et ayant été étirées grâce à leur propriété d'élasticité, ramènent le tissu à la position normale lorsque la force d'étirement n'agit plus.
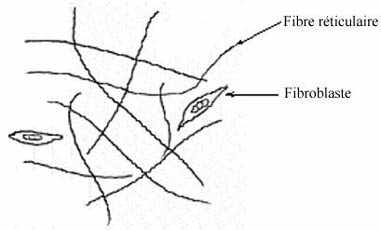
3.1.1.3 Fibres réticulaires
Les fibres réticulaires sont composées de la protéine collagène mais contiennent davantage de carbohydrates que les fibres collagéniques mêmes. Ce sont de fines fibres embranchées qui forment des réseaux délicats; elles jouent un rôle de soutien léger, par exemple en association avec des capillaires, fibres nerveuses ou musculaires, glandes et organes lymphatiques. Dans le tissu conjonctif ordinaire, elles sont synthétisées par les fibroblastes mais dans certains organes, par exemple les noeuds lymphatiques, elles le sont par d'autres cellules.
La coloration au H&E ne dévoile aucunement les fibres réticulaires. Pour ce faire, il faut utiliser l'imprégnation métallique à l'argent qui les montre noires.
3.1.2 COMPOSANTE AMORPHE
Comme son nom l'indique, la composante amorphe, ou substance amorphe, n'a pas de forme. Elle peut exister à l'état aqueux, le fluide intercellulaire, ou à l'état de gel visqueux semi-fluide. Elle provient surtout des capillaires artériels et sa composition ressemble à celle du plasma, avec des variations biochimiques locales.
La substance amorphe est riche en mucopolysaccharides, composés d'acide hyaluronique et de sucres aminés, liés en alternance et formant de longues chaînes moléculaires: les glycosaminoglycans. Ceux-ci peuvent être sulfatés ou non. Les glycosaminoglycans sulfatés sont presque toujours liés par covalence à des protéines, formant des protéoglycans: sulfates de chondroïtine A,B,C; sulfates de kératane; sulfates d'héparine.
La substance amorphe est retenue par les fibres délicates du tissu conjonctif. Les capillaires artériels, veineux et lymphatiques la traversent et contribuent à son métabolisme (ajout, retrait, modifications locales). La substance amorphe a la propension de retenir de grandes quantités d'eau. Elle constitue un milieu pour la diffusion de l'O2 et des substances nutritives, des capillaires aux cellules environnantes, de même que la diffusion inverse du CO2 et des déchets métaboliques. Une accumulation exagérée de fluide intercellulaire cause
l'oedème. Il y a généralement plus de fluide intercellulaire apporté par les capillaires artériels qu'il y en a de résorbé par les capillaires veineux; l'excès est drainé par les capillaires lymphatiques. Ceci contribue à la formation de la lymphe (du latin eau claire), dont la composition ressemble à celle du fluide intercellulaire. La substance amorphe est le site habituel des réactions d'inflammation et elle participe aux réactions immunitaires.
La quantité relative de la composante amorphe du tissu conjonctif, par rapport à la composante fibreuse, est reliée à l'âge. Abondante chez l'embryon et le jeune, la substance amorphe diminue graduellement en vieillissant. Sur la peau ce phénomène se traduit en rides.
3.2 ELEMENTS CELLULAIRES DU TISSU CONJONCTIF ORDINAIRE (voir schémas)
Les cellules retrouvées dans le tissu conjonctif ordinaire dérivent directement ou indirectement du mésenchyme, d'origine mésodermique. Certaines proviennent des cellules mésenchymateuses locales (situées au même endroit) de l'embryon, d'autres de régions éloignées mais elles migrent via le flot sanguin, pré- ou post-natalement, pour habiter le tissu conjonctif.
Le mésenchyme embryonnaire consiste en un genre de réseau lâche de cellules étoilées, à cytoplasme très peu coloré. Les cellules mésenchymateuses sont séparées les unes des autres par un gel intercellulaire renfermant de délicates fibres. Elles subissent de nombreuses mitoses et, par différenciation (revoir 1.2), sont directement ou indirectement l'ancêtre de toutes les cellules du tissu conjonctif ordinaire.
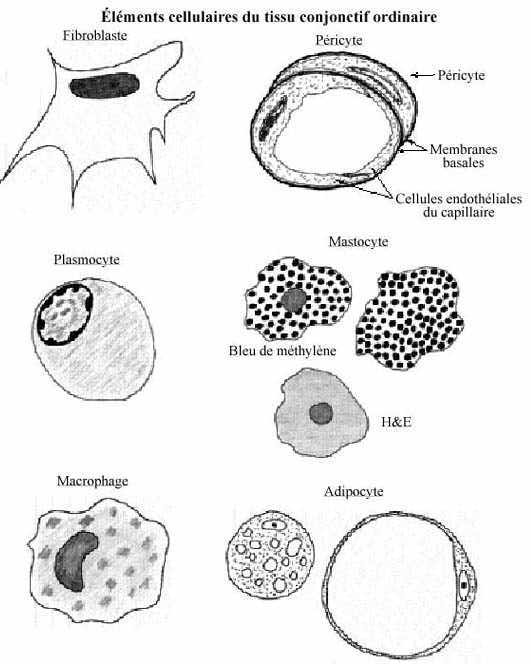
3.2.1 FIBROBLASTES
Les fibroblastes produisent les fibres du matériel intercellulaire. Ce sont les cellules les plus nombreuses du tissu conjonctif ordinaire. Au H&E, les fibroblastes ont une forme irrégulière et leur cytoplasme est tellement pâle qu'on n'en voit souvent que le noyau ovoïde. Ils prennent origine directement des cellules mésenchymateuses de l'embryon, ainsi que des péricytes (voir 3.2.2). Les fibroblastes sont des cellules sécrétrices mais, contrairement aux cellules épithéliales polarisées (revoir 2.1.3), sécrètent sur toute leur surface membranaire et deviennent ainsi enrobés dans leurs sécrétions. Leurs principales sécrétions sont le procollagène, précurseur du collagène, la proélastine, précurseur de l'élastine, et des glycosaminoglycans et protéoglycans incorporés à la substance amorphe. Les fibrocytes, de vieux fibroblastes qui ont à peu près achevé de synthétiser du matériel intercellulaire, ne se divisent probablement plus.
3.2.2 PERICYTES
Pâles cellules aplaties à la surface des capillaires, les péricytes retiennent une certaine potentialité mésenchymale. Ils peuvent être à l'origine des fibroblastes et de certains autres types cellulaires du tissu conjonctif.
3.2.3 PLASMOCYTES
Grosses cellules rondes, au noyau excentré contenant une chromatine condensée souvent périphérique, et au cytoplasme basophile abondant, les plasmocytes abondent particulièrement dans les organes lymphatiques. Dans le tissu conjonctif, ils participent aux réactions immunitaires; ils synthétisent et sécrètent des anticorps en réponse à une invasion antigénique. Ils proviennent des lymphocytes (voir 6.1.2.2.1), dont ils constituent le stade ultime de différenciation.
3.2.4 MASTOCYTES
Les mastocytes sont des cellules dont le nombre varie beaucoup selon les espèces. On les retrouve le long des petits vaisseaux sanguins, surtout les capillaires et les veinules. Leur contour est irrégulier, leur noyau rond et centré, et leur cytoplasme bourré de granules de 0.2 à 0.8µm de diamètre. Ces granules, peu évidents au H&E, obscurcissent le noyau quand colorés au bleu de méthylène. Ils contiennent de l'héparine (un glycosaminoglycans sulfaté), de l'histamine et de la sérotonine. Les mastocytes jouent un rôle dans les réactions d'allergie et d'immunité; ils sont nombreux dans le tissu conjonctif lâche sous-jacent aux épithélia tapissant les voies respiratoires et intestinales. Il semble que les mastocytes proviennent des péricytes. Ils ressemblent aux leucocytes basophiles du sang (voir 6.1.2.1.3) mais leur origine diffère.
3.2.5 MACROPHAGES (Gros mangeurs)
Les macrophages sont des cellules ovoïdes à noyau souvent excentré, ayant une chromatine plus condensée que celle du fibroblaste mais moins que celle du plasmocyte, et de taille intermédiaire entre ces deux cellules, quoique variable. Une de leurs fonctions importantes consiste à phagocyter des déchets, macromolécules, particules étrangères, vieilles cellules sanguines et bactéries. Leur propriété phagocytaire permet de les démontrer par la coloration intravitale. Ils participent aux réactions immunitaires grâce aux anticorps qu'ils possèdent à leur surface membranaire. Ils proviennent des monocytes du sang, une fois que ceux-ci gagnent le compartiment extra-sanguin (voir 6.1.2.2.2).
3.2.6 ADIPOCYTES
Les cellules adipeuses, seules ou en petits groupes, se retrouvent fréquemment dans le tissu conjonctif lâche. Les adipocytes constituent une lignée spéciale de cellules conjonctives, issues des cellules mésenchymateuses de l'embryon. Une telle cellule se différencie en adipocyte quand des gouttelettes lipidiques s'accumulent dans son cytoplasme. Quand leur nombre augmente, les gouttelettes fusionnent et peuvent former une grosse goutte unique qui remplit à peu près tout le cytoplasme de la cellule, réduisant celui-ci à un mince filet périphérique. Le diamètre d'un adipocyte peut atteindre 120µm. Les gouttelettes sont perçues comme des trous au H&E car les procédés histologiques coutumiers dissolvent la graisse. Il faut utiliser des fixateurs et procédés spéciaux pour la préserver.
L'activité mitotique des cellules destinées à devenir adipocytes est à peu près complétée peu après la naissance et les adipocytes mûrs ne se divisent plus; leur vie est très longue. Les péricytes gardent peut-être la possibilité de se différencier en adipocytes.
D'autres types cellulaires se rencontrent, en moindre quantité, dans le tissu conjonctif ordinaire, surtout aréolaire. Les leucocytes sanguins (voir 6.1.2) migrent hors des vaisseaux et sont notablement nombreux lors des réactions inflammatoires.
3.3 TISSU CONJONCTIF ORDINAIRE LACHE (AREOLAIRE) ET DENSE
Le tissu conjonctif ordinaire lâche, ou aréolaire, se compose d'une grande proportion de substance amorphe et d'éléments cellulaires, mais de relativement peu d'éléments fibreux. Ceux-ci adoptent un agencement plus ou moins bien ordonné. On y retrouve à peu près tous les types cellulaires décrits ci-haut. Souvent associé au tissu adipeux, le tissu conjonctif lâche se rencontre sous le derme de la peau, autour des vaisseaux sanguins et nerfs, et remplit les interstices entre les organes.
Le tissu conjonctif ordinaire dense contient une plus grande proportion d'éléments fibreux. Il est dit régulier dans le derme de la peau, où les fibres collagéniques sont sinueuses et parallèles, et dans les tendons et ligaments, où elles sont droites et parallèles. Les tendons et ligaments ne renferment qu'un type cellulaire, les fibroblastes, et peu ou pas de vaisseaux sanguins. Ce tissu conjonctif dense et régulier, doté de résistance considérable, peut subir de grandes forces de tension dans le sens des fibres sans être étiré ou déchiré, comme il se doit des tendons et ligaments rattachant les muscles aux os ou les os entre eux. Les ligaments élastiques consistent davantage en élastine qu'en collagène.
Le tissu conjonctif dense peut être irrégulier ou enchevêtré, les fibres collagéniques orientées dans plusieurs directions mais pouvant former un feuillet, comme dans les capsules recouvrant les viscères, le fascia recouvrant les muscles ou la dure-mère recouvrant le système nerveux central. Ce tissu peut subir des tensions dans toutes les directions sans se briser, protégeant ainsi l'organe recouvert.
3.4 TISSU ADIPEUX
Le tissu conjonctif presque entièrement composé de cellules adipeuses s'appelle tissu adipeux. Les adipocytes y sont organisés en groupes appelés lobules, séparés par des partitions (septa) de tissu conjonctif délicat, collagénique et réticulaire, infiltrées de vaisseaux sanguins et nerfs. Il existe deux types principaux de tissu adipeux: blanc et brun.
3.4.1 TISSU ADIPEUX BLANC
Le plus retrouvé chez les mammifères, le tissu adipeux blanc est en réalité jaunâtre à cause de sa teneur en carotène, un pigment. Chez l'humain, il constitue de 15 à 20% du poids corporel chez l'homme, 20 à 25% chez la femme. C'est un tissu métaboliquement très actif, notamment dans l'absorption, la synthèse, la mise en réserve et la mobilisation de lipides neutres. A la température du corps, la graisse se trouve sous forme d'huile. Elle se compose de triglycérides qui constituent une réserve énergétique à haute teneur calorifique. Le métabolisme de la graisse est grandement influencé par les hormones et le système nerveux. Le tissu adipeux blanc remplit les interstices entre les organes, minimisant la friction entre eux, et agit comme amortisseur (fesses, paumes des mains, plantes des pieds). Il sert aussi d'isolant thermique pour les habitants des pays froids.
3.4.2 TISSU ADIPEUX BRUN
Le tissu adipeux brun assure la régulation de la température corporelle chez le nouveau-né puis disparaît chez plusieurs espèces. Chez l'adulte, il se rencontre surtout chez les mammifères qui hibernent, chez qui il constitue une source de chaleur au sortir de l'hibernation. Sa couleur brunâtre révèle sa riche vascularisation et la teneur élevée des adipocytes en mitochondries (cytochrome oxydase).
{kind=link}
{kind=link}
{kind=link}