> Sang
> Tissu myéloïde et hémopoïèse
CHAPITRE 6: SANG, TISSU MYELOIDE ET HEMOPOIESE
6.1 SANG
Le sang constitue une classe spéciale de tissu conjonctif. Les éléments cellulaires, les cellules sanguines, ne sont pas rattachées les unes aux autres, ni ne sont retenues par du matériel intercellulaire, comme c'est le cas de la plupart des autres cellules du tissu conjonctif. Elles appartiennent à une catégorie de cellules libres. En suspension dans le plasma, elles sont transportées par lui. Le plasma représente le matériel intercellulaire, fluide, du sang. Les cellules sanguines sont formées dans les organes hémopoïétiques, à partir desquels elles gagnent la circulation. D'autres particules se retrouvent en suspension dans le plasma: plaquettes, globules lipidiques et hémaconies ou "poussières sanguines": produits de désintégration des vieilles cellules sanguines.
Le sang est l'un des tissus les plus faciles à étudier, même chez l'humain, car il peut être prélevé aisément par véniponcture ou, encore plus simplement, en piquant le bout du doigt et en étalant une goutte sur une lame; ce frottis frais peut être examiné immédiatement mais, de préférence, on le laisse sécher, on le colore puis le recouvre d'une lamelle: le frottis sec. Plusieurs techniques de coloration existent pour mettre en évidence les différentes cellules sanguines, mais la plus coutumière est celle de Wright-Giemsa, utilisant de l'éosine jaunâtre, du bleu de méthylène, de l'azur A, de l'azur B et du violet de méthylène.
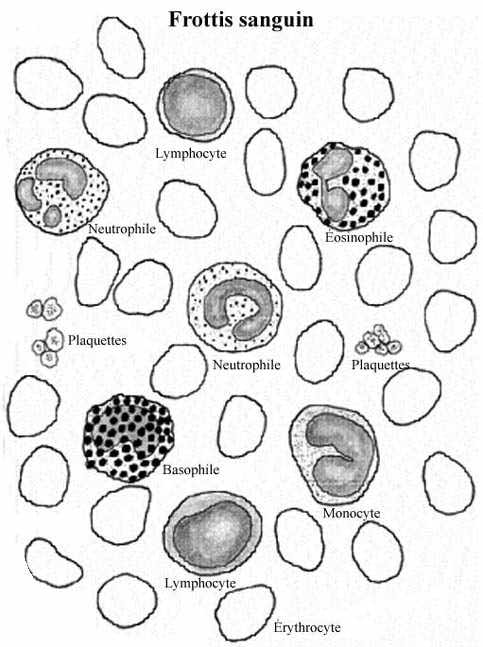
On reconnaît deux grandes catégories de cellules sanguines, les "globules" rouges: érythrocytes ou hématies, et les "globules" blancs: leucocytes.
6.1.1 ERYTHROCYTES (GLOBULES ROUGES, HEMATIES)
Les érythrocytes sont de 500 à 1000 fois plus nombreux que les leucocytes: 5 X 106/cc de sang.
Chez les mammifères, l'érythrocyte mûr ne possède pas de noyau. Celui-ci a été expulsé en fin de maturation, dans le tissu hémopoïétique, avant que l'érythrocyte n'entre en circulation. L'érythrocyte anucléé adopte plus ou moins la forme d'un disque biconcave, notamment chez l'humain. Les constituantes moléculaires particulières de sa membrane cellulaire et la composition du complexe colloïdal qui tient lieu de cytoplasme déterminent la forme particulière de l'érythrocyte. Elles lui confèrent flexibilité et élasticité qui lui permettent de subir les déformations nécessaires pour circuler dans les vaisseaux sanguins de faible calibre.
Sur frottis, le plus grand diamètre d'un érythrocyte anucléé (disque biconcave), 7.2µm, varie très peu en condition physiologique. Ce critère fiable peut donc s'employer pour juger de pathologies sanguines possibles.
L'eau constitue 66% du contenu de l'érythrocyte et l'hémoglobine (Hb) 33%. L'Hb se compose de la protéine globine et du pigment hème qui contient du fer et qui confère la coloration rouge au complexe. D'autres protéines et enzymes, notamment prothrombine et fibrinogène, ainsi que des lipides contribuent à la formation du complexe colloïdal.
Dans la circulation pulmonaire, l'érythrocyte, grâce à l'Hb, lie l'O2 de l'air inspiré et le livre aux autres tissus par la circulation systémique (voir Chapitre 10). Dans les tissus, l'Hb désoxygénée (réduite) lie le CO2 pour éventuellement le larguer dans l'air expiré. Il est important que l'interface entre l'érythrocyte et le plasma soit la plus grande possible pour maximiser la diffusion des gaz respiratoires. Sa forme aplatie minimise la distance cytoplasmique de diffusion. L'avantage de l'état anucléé est que toute la cellule peut contenir de l'Hb et, ainsi, accroître son efficacité par unité de volume. Le prix à payer pour l'absence de noyau est une durée de vie courte. Effectivement, la vie de l'érythrocyte se limite à quelques mois, après quoi il dégénère.
Chez les vertébrés sous-mammaliens, l'érythrocyte conserve son noyau et par conséquent une forme généralement arrondie.
6.1.2 LEUCOCYTES (GLOBULES BLANCS)
A l'instar des érythrocytes, les leucocytes sont formés dans les organes hémopoïétiques puis gagnent la circulation sanguine. Contrairement aux érythrocytes qui remplissent leurs fonctions dans le sang même, les leucocytes quittent la circulation et accomplissent leurs tâches une fois rendus dans le tissu conjonctif. Donc, les leucocytes sont des cellules sanguines parce qu'ils empruntent la circulation sanguine comme moyen de transport, mais ils n'y accomplissent pas leurs fonctions.
Sur frottis, on distingue 5 types de leucocytes, appartenant à deux groupes principaux: granulocytes et agranulocytes. Tous conservent leur noyau à l'état mûr. La distinction entre granulocytes et agranulocytes repose sur le fait que ceux-là possèdent des granules cytoplasmiques bien visibles tandis que ceux-ci n'en possèdent pas, du moins en microscopie photonique, ou ils sont si fins qu'à toute fin pratique invisibles.
6.1.2.1 Granulocytes
On décrit 3 types de leucocytes franchement granulocytaires:
éosinophile: dont les granules se colorent bien aux colorants acides, notamment l'éosine.
basophile: dont les granules se colorent bien aux colorants basiques.
neutrophile: dont les granules se colorent bien sans présenter d'affinité particulière pour l'un ou l'autre type de colorant à pH neutre.
Ayant un noyau ultilobé pouvant présenter plusieurs formes, les granulocytes portent aussi le nom de polymorphonucléaires, ou polymorphes, qualifiés selon leur propriété tinctoriale: polymorphonucléaire éosinophile (PME), basophile (PMB) et neutrophile (PMN).
6.1.2.1.1 Neutrophiles (PMN)
Les neutrophiles constituent de 50 à 70% des leucocytes (granulocytes et agranulocytes ensemble): 3 à 6 X 103/cc de sang.
Le diamètre du neutrophile atteint 10 à 12µm. A cause de son noyau multilobé, 2 à 5 lobes reliés par de minces filets nucléaires, il peut donner l'impression d'être multinucléé car les minces filets ne sont pas toujours visibles. La chromatine grossière et condensée occupe une position périphérique, en bordure de la membrane nucléaire; elle se colore intensément aux colorants basiques. Le cytoplasme occupe un plus grand volume cellulaire que le noyau (taux nucléoplasmique faible) et renferme deux sortes de granules: granules neutrophiles ou spécifiques, si fins que difficilement visibles, et granules azurophiles, plus gros et rouge pourpre. Les granules neutrophiles renferment un bactéricide et l'enzyme phosphatase alcaline. Les granules azurophiles représentent des lysosomes qui contiennent au moins 6 enzymes hydrolytiques. Les PMN ne séjournent que quelques heures dans la circulation sanguine et pénètrent dans le tissu conjonctif lâche, où ils entrent en jeu dans le processus d'inflammation aiguë; ils s'attaquent particulièrement aux bactéries.
6.1.2.1.2 Eosinophiles (PME)
Les éosinophiles constituent de 1 à 4% des leucocytes: 120-350/cc de sang, et leur nombre fluctue, notamment avec le cycle diurne: plus nombreux la nuit et moins le matin.
Le diamètre de l'éosinophile atteint 12 à 17µm. Son noyau bilobé contient une chromatine périphérique moins condensée et moins colorée que celle du neutrophile. Son cytoplasme est bourré de gros granules réfractaires rouges ou orangés, riches en peroxydases et autres enzymes; ce sont des lysosomes. Des petits granules contiennent de l'arylsulfatase et de la phosphatase acide. Abondants dans le tissu conjonctif de l'intestin, des poumons, de la peau et des parties génitales externes, les éosinophiles participent aux réactions d'allergie et phagocytent les complexes antigènes-anticorps libres.
6.1.2.1.3 Basophiles (PMB)
Les basophiles constituent 0.5% des leucocytes: 40/cc de sang.
La taille du basophile est la même que celle du neutrophile (10-12µm) mais son taux nucléoplasmique est plus élevé. Le noyau, bilobé ou segmenté et de forme irrégulière, se colore moins intensément que celui des autres granulocytes. De gros (0.5µm) granules cytoplasmiques bleu foncé, renfermant de l'héparine, l'obscurcissent souvent. De plus, les basophiles contiennent environ la moitié de l'histamine détectée dans le sang. Ils s'accumulent aux sites d'inflammation et interviennent dans les réactions d'allergie.
6.1.2.2 Agranulocytes
6.1.2.2.1 Lymphocytes
Après les neutrophiles, les lymphocytes constituent le deuxième type leucocytaire le plus nombreux, soit 20 à 40% des leucocytes: 1000 à 4000/cc de sang.
Le lymphocyte est la seule cellule sanguine à se trouver également dans la circulation lymphatique. Sans spécialisation morphologique remarquable, on parle de lymphocyte petit, d'un diamètre 7 à 8µm, ou de lymphocyte moyen, d'environ 12 µm. De gros lymphocytes résident dans les organes lymphatiques. Le lymphocyte possède un noyau parfois incurvé et à chromatine très condensée, puis un cytoplasme peu abondant, à basophilie variable, contenant quelques granules azurophiles (lysosomes). Le lymphocyte se déplace très rapidement et participe aux réactions immunitaires. Il se différencie en plasmocyte
6.1.2.2.2 Monocytes
Les monocytes constituent de 2 à 8% des leucocytes : 200 à 600/cc de sang.
Le monocyte peut ressembler au lymphocyte moyen ou au métamyélocyte neutrophile (voir 6.2.2.2). Avec un diamètre de 12 à 18µm, c'est le leucocyte le plus gros. Il adopte une forme plus ou moins sphérique mais la forme de son noyau varie d'ovoïde à légèrement invaginée ou même franchement en fer à cheval. La chromatine périphérique, moins dense que celle du lymphocyte, se colore bleu violet. Le cytoplasme gris bleu occupe une bonne proportion du volume cellulaire; il est dépourvu de granules spécifiques mais non de granules azurophiles (lysosomes). Plus que toute autre cellule sanguine, le monocyte se caractérise par sa propriété de motilité, qui se traduit par l'existence de nombreux pseudopodes. Le monocyte est le précurseur immédiat du macrophage (revoir 3.2.5). Ainsi, les cellules de cette lignée portent le nom de monocytes dans la circulation sanguine et de macrophages dans le tissu conjonctif.
6.1.3 PLAQUETTES
Les plaquettes sont de petits fragments cytoplasmiques détachés de très grosses cellules de la moelle osseuse, les mégacaryocytes. Disque ovale biconvexe ayant une région centrale granulaire, le granulomère, et une région externe bleu pâle, l'hyalomère, chaque plaquette est délimitée par une membrane cytoplasmique. Leur nombre varie entre 250 et 350x103/cc de sang. Les plaquettes, isolées en suspension dans le plasma, s'agrègent quand stimulées et adhèrent à la paroi interne d'un vaisseau endommagé pour en endiguer l'hémorragie. Cet événement s'accompagne généralement de la formation de la protéine fibreuse fibrine, dérivée du précurseur plasmatique fibrinogène, complétant le phénomène de coagulation. Ces mêmes processus d'agrégation, d'adhésion et de coagulation avec l'âge peuvent causer l'artériosclérose.
Chez les vertébrés sous-mammaliens, de petites cellules nucléées appelées thrombocytes jouent le même rôle que les plaquettes. On appelait d'ailleurs les plaquettes des thrombocytes, mais ce terme devrait être réservé aux cellules nucléées ne provenant pas des mégacaryocytes.
6.2 TISSU MYELOIDE ET HEMOPOIESE
Hémopoïèse (hématopoïèse) signifie formation des cellules sanguines, un processus en cours pendant toute la vie de l'animal. Elle a lieu dans les organes hémopoïétiques. Chez l'embryon, l'hémopoïèse se produit d'abord dans la paroi du sac vitellin, puis dans le foie, la rate et la moelle osseuse. Postnatalement, la moelle osseuse et la rate deviennent les sites principaux d'hémopoïèse, puis la moelle osseuse seule chez certaines espèces. Chez l'embryon, l'hémopoïèse se produit également à partir de cellules mésenchymateuses retrouvées dans la lumière des capillaires en formation.
Les premières cellules à caractère hémopoïétique sont des cellules pluripotentes, pouvant donner naissance à toutes les catégories de cellules sanguines, de même qu'aux cellules endothéliales bordant les vaisseaux sanguins (voir Chapitre 10) et aux cellules à caractère macrophagique comme les cellules microgliales du système nerveux ou les cellules de Kupffer du foie. Ces cellules pluripotentes hémopoïétiques se regroupent en îlots sanguins.
Chez le mammifère adulte, érythrocytes, leucocytes et plaquettes sont surtout formés dans la moelle osseuse. Les lymphocytes subissent leur maturation dans les organes lymphatiques: thymus, noeuds lymphatiques et rate, mais leurs cellules-souche proviennent de la moelle osseuse. Dans ce chapitre-ci, nous étudierons la moelle osseuse et l'hémopoïèse qui s'y produit.
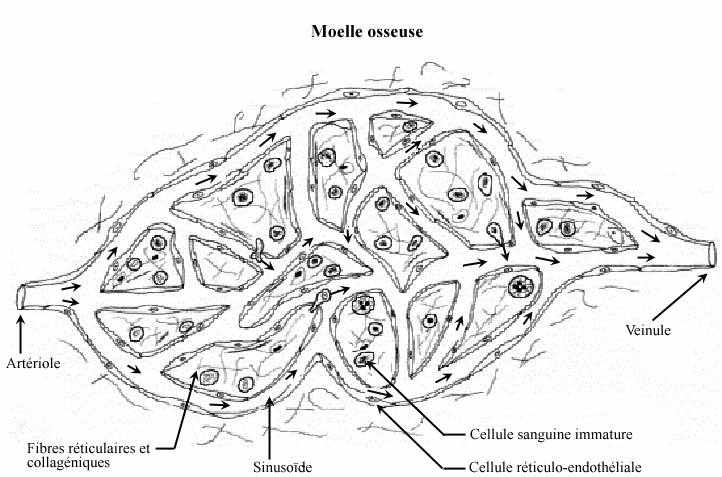
6.2.1 MOELLE OSSEUSE OU TISSU MYELOIDE (grec: myelos= moelle)
La moelle osseuse est reliée à un circuit vasculaire: une artère traverse la paroi de l'os et pénètre dans la cavité, s'embranche en artérioles qui se ramifient elles aussi, et les petits vaisseaux du réseau se réunissent éventuellement en veinules puis en une veine qui quitte la cavité et ressort de l'os. Les ramifications sont analogues aux réseaux capillaires traversant n'importe quel autre organe sauf que, plutôt que de parler de capillaires (voir 10.3), on appelle les petits vaisseaux de la moelle osseuse des sinusoïdes. Ainsi, la moelle osseuse est constituée d'une multitude de sinusoïdes retenus par un réseau tridimensionnel de fibres réticulaires (revoir 3.1.1.3) et de fines fibres de collagène. Les interstices entre les fibres contiennent de nombreuses cellules de la lignée sanguine à différents stades de différenciation. Une fois mûres, les cellules sanguines: érythrocytes et les 5 types leucocytaires, en plus des plaquettes, traversent la paroi des sinusoïdes en empruntant des pores de migration labiles, par un mécanisme actif appelé diapédèse. Dans la lumière des sinusoïdes, elles se trouvent à entrer en circulation et sont amenées hors de la moelle.
Outre les cellules sanguines, dont les étapes de différenciation seront bientôt décrites, la moelle osseuse renferme d'autres éléments cellulaires:
Les cellules réticulo-endothéliales, comme le terme "endothéliales" laisse deviner, constituent, à elles seules, la paroi des sinusoïdes: ces capillaires à très mince paroi mais à lumière agrandie. A cause de l'extrême minceur et de la très faible coloration des cellules les constituant, les sinusoïdes sont rarement perçus en préparation ordinaire. En plus de leur fonction structurale, les cellules réticulo-endothéliales sécrètent les fibres réticulaires, comme le préfixe "réticulo" le suppose. Les fibres réticulaires offrent un substrat anatomique aux cellules hémopoïétiques en formation.
Les fibroblastes élaborent les fibres collagéniques qui supportent les artérioles, sinusoïdes et veinules de la moelle osseuse.
Les macrophages, nombreux dans la moelle osseuse, phagocytent les cellules sanguines et les plaquettes défectueuses, en plus de participer à l'induction des cellules-souche hémopoïétiques.
Les adipocytes sont en quantité variable, selon la moelle (voir plus bas).
Les cellules ostéogéniques, cellules-souche des ostéoblastes, retrouvées en bordure de la cavité osseuse, accomplissent une fonction additionnelle: l'induction des cellules hémopoïétiques à se différencier. Réciproquement, les cellules-souche hémopoïétiques jouent un rôle d'inducteur sur les cellules ostéogéniques. Il en ressort que les fonctions ostéogénique et hémopoïétique agissent l'une sur l'autre, selon des mécanismes compliqués et mal compris.
Chez le foetus, presque toute la moelle osseuse est rouge, en activité intense d'hémopoïèse. La couleur de la moelle rouge reflète sa grande activité hémopoïétique et, de ce fait, sa haute teneur en érythrocytes. Dans plusieurs os, au cours du vieillissement, la moelle rouge se transforme graduellement en moelle jaune, qui ne produit plus de cellules sanguines. Le tissu adipeux remplace le tissu hémopoïétique. Chez l'adulte, la moelle rouge ne se retrouve plus que dans la diploë des os crâniens, les côtes, le sternum, les vertèbres, certains os courts et les épiphyses. Dans des cas de stress extrême et de famine, la moelle jaune peut se transformer en moelle rouge.
6.2.2 HEMOPOIESE (Hématopoïèse)
Comme pour les cellules sanguines en circulation, l'étude de l'hémopoïèse s'effectue par frottis de moelle, généralement obtenu par ponction sternale, frottis étalé sur lame, séché et coloré au Wright-Giemsa. Inutile de préciser que, sur frottis, l'organisation structurale du stroma conjonctif est perdue, de même que l'intégrité des sinusoïdes.
Au cours du développement embryonnaire, les premières cellules à se différencier sont celles de la lignée érythrocytaire. C'est ensuite que les lignées leucocytaires se manifestent. Chez l'adulte, la production des différentes lignées de cellules sanguines s'effectue selon la proportion relative normale des cellules mûres mais également en réponse à des besoins ponctuels de l'organisme: la fatigue, une invasion microbienne ou virale, une allergie, etc.
Les différentes cellules sanguines prennent toutes origine d'une cellule-souche pluripotente, l'unité de formation de colonie ou CFU (colony forming unit). Petites cellules libres de 7 à 10µm de diamètre, les CFU sont peu nombreux et non identifiables en préparation ordinaire.
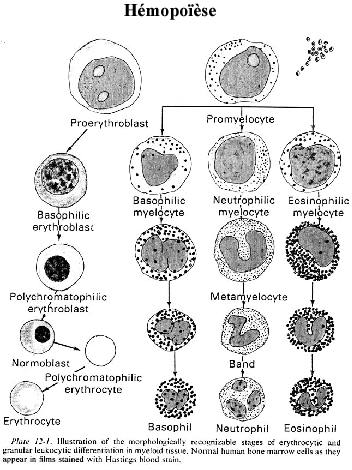
6.2.2.1 Erythropoïèse
La production d'érythrocytes est stimulée par l'hypoxie (baisse d'O2), via l'hormone érythropoïétine. Cette hormone, fabriquée dans le rein, induit les CFU qui lui sont sensibles à se différencier sur la lignée érythrocytaire. La première cellule de cette lignée facilement reconnaissable au microscope est le proérythroblaste. Cellule de 12 à 15µm de diamètre, son noyau contient une chromatine finement granulaire et son cytoplasme est légèrement basophile.
Le proérythroblaste se divise et se différencie en érythroblaste basophile, quelque peu plus petit que son prédécesseur. Son noyau arrondi contient une chromatine plus condensée et la basophilie de son cytoplasme s'attribue à son contenu élevé en polyribosomes.
L'érythroblaste basophile se divise et se différencie en érythroblaste polychromatophile, à noyau plus petit renfermant une chromatine encore plus condensée, sous forme de gros granules, et à cytoplasme bleu rosé. La qualité polychromatophile de son cytoplasme tient des polyribosomes basophiles et de l'hémoglobine acidophile nouvellement synthétisée. L'érythroblaste polychromatophile est peut-être le dernier de la lignée à pouvoir se diviser.
Il se différencie en érythrocyte polychromatophile, aussi appelé réticulocyte (terme à éviter), à cytoplasme polychromatophile et dont le noyau commence à être expulsé. Chez les mammifères, l'érythrocyte polychromatophile se différencie davantage en perdant sa basophilie cytoplasmique et en continuant d'expulser son noyau; c'est l'érythrocyte orthochromatique ou normoblaste. Le normoblaste expulse complètement son noyau, c'est l'érythrocyte normocytaire ou simplement érythrocyte mûr. Les macrophages du stroma phagocytent les noyaux expulsés. L'érythrocyte peut maintenant entrer en circulation, comme décrit plus haut.
6.2.2.2 Granulocytopoïèse
La formation des leucocytes granulaires est stimulée par l'hormone leucopoïétine, à partir de CFU qui lui sont sensibles. La première cellule de cette lignée facilement reconnaissable au microscope est le promyélocyte, contenant quelques granules cytoplasmiques non encore attribuables à l'un ou l'autre des trois types granulocytaires. Le promyélocyte se divise et se différencie en myélocyte, de taille réduite, à noyau ovale ou même échancré, et à cytoplasme granulaire.
Le myélocyte se différencie en métamyélocyte contenant des granules identifiables: métamyélocyte neutrophile, métamyélocyte éosinophile, métamyélocyte basophile. Chacun des trois types acquiert davantage de granules caractéristiques et continue sa différenciation en cellule mûre, notamment lobulation du noyau et condensation de la chromatine à la périphérie. Les cellules mûres gagnent la circulation sanguine.
6.2.2.3 Monocytopoïèse
Dérivant toujours des CFU, le premier stade de différenciation identifiable est le promonocyte ou monoblaste, semblable à un petit lymphocyte. Le promonocyte se divise et se différencie en monocyte qui gagne la circulation sanguine. Peu après, il pénètre les tissus en tant que macrophage.
6.2.2.4 Lymphocytopoïèse
Plusieurs lymphocytes sont formés dans le tissu myéloïde, à partir de CFU, mais leurs stades développementaux se reconnaissent difficilement. Ils migrent souvent à des stades précoces vers les organes lymphatiques où ils terminent leur maturation. Il semble que celle-ci soit davantage biochimique que morphologique, du moins en ce qui peut être jugé au microscope photonique.
Il faut noter que la terminologie de l'hémopoïèse utilisée dans ici est relativement simple et peu contestable. Il faut toutefois être conscient qu'il existe à peu près autant de stades développementaux et de termes pour les décrire qu'il existe d'auteurs.
6.2.2.5 Mégacaryocytopoïèse et formation des plaquettes
Provenant aussi de CFU, le mégacaryocyte est la plus grosse cellule observée dans la moelle osseuse. Il a subi de nombreuses mitoses sans distribuer ses chromosomes-fils dans des noyaux différents, créant un noyau multilobé polyploïde. Le cytoplasme est partitionné par un système de membrane anastomosée. La taille de chaque compartiment égale à peu près celle d'une plaquette. Les compartiments cytoplasmiques, délimités de membrane, se détachent et traversent la paroi des sinusoïdes pour s'incorporer à la circulation. Quand toutes les plaquettes ont été détachées d'un mégacaryocyte, le noyau et le peu de cytoplasme restants sont phagocytés par les macrophages.
{kind=link}
{kind=link}
{kind=link}