CHAPITRE 7: TISSU MUSCULAIRE
C'est chez la cellule musculaire, ou myocyte, que la contractilité, propriété fondamentale de toute cellule animale, atteint sa plus grande expression. Presque tout le cytoplasme du myocyte consiste en machinerie contractile. Les cellules musculaires étant longues et minces, on les appelle souvent fibres musculaires. Elles ne sont néanmoins pas des composantes extracellulaires comme les fibres de collagène.Le muscle s'accompagne d'une bonne quantité de tissu conjonctif dans lequel reposent nerfs et vaisseaux sanguins le desservant. Vu son travail vigoureux, l'apport en O2 et en substances nutritives est considérable. Les besoins contractiles varient selon les régions de l'organisme et selon les organes et, pour un organe donné, ces besoins varient dans le temps. C'est ainsi qu'il existe trois types de tissu musculaire, reliés à des fonctions différentes, les trois d'origine mésodermique. Chaque type musculaire possède son type de myocyte.
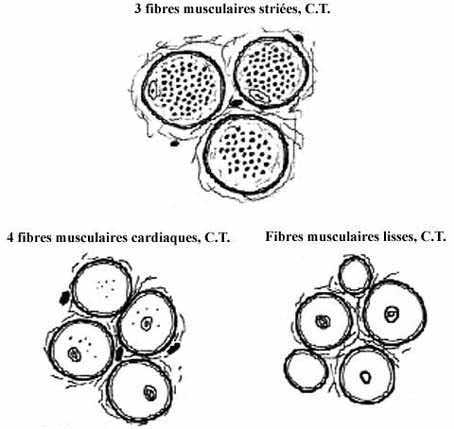
Voir les schémas sur les fibres musculaires de différents types
7.1 MUSCLE STRIE SQUELETTIQUE
Les muscles striés squelettiques portent ce nom parce qu'ils sont rattachés, du moins par une extrémité, à une partie quelconque du squelette osseux ou cartilagineux et qu'en coupe longitudinale, leurs fibres présentent des striations régulières. Leur fonction est contrôlée volontairement; ils peuvent être contractés ou relâchés à volonté, mais fonctionnent aussi sans le contrôle volontaire, de façon réflexe. Ils sont sous la dépendance du système nerveux somatique.
Le muscle entier, i.e. l'organe (ex., biceps, gastrocnemius), est recouvert d'une solide capsule conjonctive (revoir 3.3): l'épimysium. Continus avec celui-ci, des septa de tissu conjonctif un peu moins dense divisent le muscle en compartiments et constituent le périmysium. Continu avec ce dernier, l'endomysium consiste en tissu conjonctif délicat qui entoure chaque fibre musculaire. Epimysium, périmysium et endomysium enrobent et conduisent les vaisseaux sanguins et lymphatiques ainsi que les nerfs qui desservent le muscle. A certains endroits, tissus musculaire et conjonctif se mélangent pour s'ancrer à l'os (périoste) ou au cartilage (périchondre) sur lequel ils tirent pour effectuer des mouvements. Pour ce faire, le tissu conjonctif peut former de véritables tendons.
Les fibres du muscle strié squelettique sont très grosses et multinucléées, résultant de la fusion de plusieurs myocytes qui ne se divisent plus. Leur diamètre, constant sur toute la longueur, peut atteindre 100µm et leur longueur de 1 à 40mm. Chez l'embryon, les myoblastes sont uninucléés. Plusieurs myoblastes fusionnent bout à bout et forment une fibre allongée et multinucléée: le myotube. Les myotubes développent des myofibrilles (voir plus bas) et deviennent les fibres musculaires striées. Il n'est donc pas tout à fait juste d'équivaloir fibre musculaire striée à "cellule" musculaire striée.
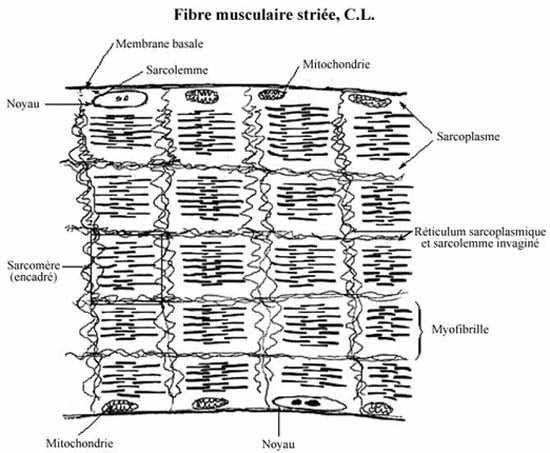
La fibre musculaire striée est délimitée par une membrane cellulaire appelée sarcolemme (grec: sarkos= chair; lemma= écorce), elle-même enveloppée d'une membrane basale, évidente au PAS. Les noyaux allongés se situent dans la région périphérique du sarcoplasme (cytoplasme), parallèles au sarcolemme. Extérieurs au sarcolemme et à sa membrane basale, se trouvent les noyaux allongés des fibroblastes de l'endomysium, qu'il faut éviter de confondre avec ceux de la fibre musculaire même.
Le sarcolemme démontre une propriété de conduction semblable à celle du neurolemme (voir Chapitre 8) et joue un rôle dans la conduction de la vague de dépolarisation qui lui est transmise à la jonction neuromusculaire. Le sarcolemme s'invagine profondément dans le sarcoplasme, créant un système de tubules à intervalles assez réguliers le long de la fibre. Ces tubules s'embranchent à l'intérieur du sarcoplasme de façon à entourer chaque unité de contraction ou sarcomère (voir plus bas). Ce système élaboré d'invaginations membranaires permet la transmission presque instantanée d'une vague de dépolarisation partout dans la fibre musculaire et assure une contraction rapide et efficace.
Le réticulum sarcoplasmique (endoplasmique) lisse consiste en un système élaboré de lamelles interconnectées qui entrent en communication avec le système tubulaire du sarcolemme. Réservoir de Ca++, ion essentiel à la contraction musculaire, le réticulum sarcoplasmique en règle la concentration dans les myofibrilles. Les mitochondries, centrales énergétiques de la cellule, sont nombreuses dans la fibre musculaire striée.
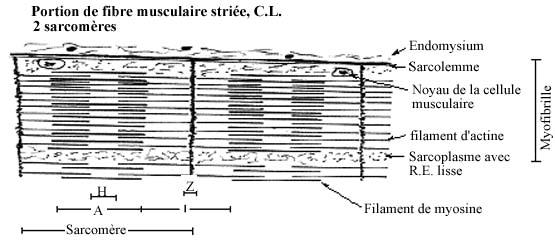
En coupe longitudinale, des stries verticales, en alternance claires et foncées, se voient le long de la fibre. Observées au microscope en lumière polarisante, les stries foncées ne laissent pas passer la lumière mais la reflètent: elles sont biréfringentes ou anisotropes, d'où leur nom de bandes A. Les stries claires laissent passer la lumière et sont dites isotropes, d'où leur nom de bandes I. Chaque bande I est traversée à son tour par une ligne foncée, appelée bande Z. Selon l'état de contraction, on peut voir une ligne pâle traversant la bande A, c'est la bande H. Ces striations sont causées par l'agencement particulier des protéines contractiles dans la fibre, et la bande Z par les tubules de sarcolemme et de réticulum sarcoplasmique traversant la fibre de part en part.
Chaque fibre musculaire striée peut être dissociée en ses fibrilles composantes, les myofibrilles, composées de protéines contractiles orientées longitudinalement; les myofibrilles portent donc le même patron de striation que la fibre entière. En coupe transversale du muscle, les myofibrilles apparaissent comme des points dans le sarcoplasme. Les myofibrilles sont délimitées par les tubules longitudinaux de sarcolemme et de réticulum sarcoplasmique (perpendiculaires à ceux de la bande Z).
On appelle sarcomère la portion d'une myofibrille entre deux bandes Z consécutives; il s'étend sur 2 à 3µm. Le sarcomère représente l'unité contractile, fonctionnelle, du muscle strié. Une contraction du muscle correspond à un raccourcissement du sarcomère.
Les myofibrilles se composent surtout de deux protéines filamenteuses, l'actine et la myosine, arrangées longitudinalement. En réponse à une vague de dépolarisation du sarcolemme, l'actine et la myosine glissent le long l'une de l'autre et se polymérisent en actomyosine, produisant un raccourcissement qui est à la base de la contraction musculaire. Ce processus est dépendant de l'ATP et du Ca++.
Étant aussi différenciées qu'elles le sont, les fibres musculaires striées squelettiques sont post-mitotiques, i.e., elles ont perdu la capacité de se diviser mitotiquement.
7.2 MUSCLE CARDIAQUE
Il constitue la plus grande partie du coeur et se retrouve dans la paroi de la veine cave inférieure à sa jonction avec l'oreillette droite. La fonction des fibres cardiaques, non sous contrôle volontaire, relève du système nerveux viscéral. Le tissu conjonctif se retrouve en quantité réduite dans le muscle cardiaque. Entre les cellules, on rencontre des capillaires, lymphatiques et nerfs.
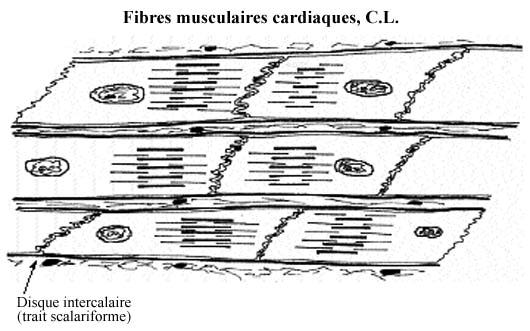
Les fibres musculaires cardiaques résultent de la fusion de quelques cardiomyocytes mais sont beaucoup plus courtes que les fibres musculaires striées squelettiques: quelques centaines de mm. Elles sont reliées bout à bout par des jonctions de types adherens et nexus. Les membranes cytoplasmiques ainsi jointes décrivent d'innombrables interdigitations, de sorte que la limite entre deux fibres est épaisse et apparaît, au microscope photonique, comme un zigzag oblique: le disque intercalaire ou trait scalariforme, typique du muscle cardiaque. Les jonctions servent une fonction mécanique mais aussi une fonction de transmission électrique.
Les fibres musculaires cardiaques possèdent un ou quelques noyaux en position plus ou moins centrale. Les stries correspondant aux bandes A, I, Z et H s'y retrouvent mais sont moins évidentes que dans la fibre striée squelettique; c'est pourquoi on ne les appelle généralement pas fibres striées. Néanmoins, il n'est pas inutile de qualifier le plus possible la fibre musculaire dont on parle pour éviter toute confusion.
Les vieux cardiomyocytes renferment souvent des granules de lipofuchsine, dérivés de lysosomes, granules retrouvés fréquemment dans les types cellulaires ayant perdu la propriété de se diviser.
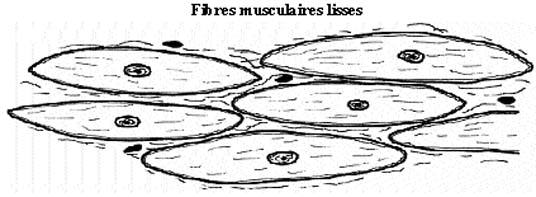
7.3 MUSCLE LISSE
Ni strié, ni volontaire, le muscle lisse subit le contrôle du système nerveux viscéral. Ses cellules tendent à s'organiser en feuillets qui contribuent à former la paroi de structures tubulaires: vaisseaux sanguins, voies respiratoires, tube digestif, etc., où leur rôle consiste à en régler le diamètre par un ajustement de leur degré de contraction. Les couches musculaires lisses peuvent néanmoins être subdivisées en faisceaux enrobés de tissu conjonctif équivalent au périmysium, mais qui n'en porte pas le nom, septa qui guident les vaisseaux sanguins et fibres nerveuses. Des fibres musculaires lisses, seules ou regroupées, se trouvent à divers autres endroits de l'organisme, comme dans le tissu conjonctif lâche, le derme de la peau, etc.
Fusiforme, la cellule musculaire lisse a une taille variable: longueur de 30µm dans les artérioles et jusqu'à 500µm dans l'utérus gravide; moyenne de 8µm de diamètre central et 200µm de long. Le noyau unique et central s'entoure de cytoplasme d'apparence lisse. Bien que non strié à proprement parler, ce type de myocyte renferme néanmoins, dans la périphérie du cytoplasme, des filaments d'actine et de myosine qui se polymérisent et induisent la contraction, moins spectaculaire que celle du muscle strié. Moins spécialisé que les deux autres types cellulaires, le myocyte lisse conserve la propriété de se diviser.
Il peut parfois être difficile de différencier muscle lisse et tissu conjonctif sur une coupe colorée au H&E. La taille des éléments et la position des noyaux s'avèrent des critères utiles de distinction.
7.4 CELLULES MYOEPITHELIALES ET CELLULES MYOIDES
Deux types de cellules ressemblent morphologiquement aux cellules musculaires lisses et se montrent capables de contractions sous l'influence d'hormones ou d'influx nerveux. Les cellules myoépithéliales se rencontrent autour des unités sécrétrices des glandes exocrines (revoir 2.2.1). Elles dérivent de l'ectoderme embryonnaire plutôt que du mésoderme. Les cellules myoïdes ou péritubulaires entourent les tubules séminifères du testicule (voir 16.1). Dans les deux cas, la contraction des cellules favorise l'excrétion ou la circulation des produits contenus dans les structures qu'elles entourent.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}