> Neurones
> Système nerveux périphérique
> Synapses
CHAPITRE 8: TISSU NERVEUX
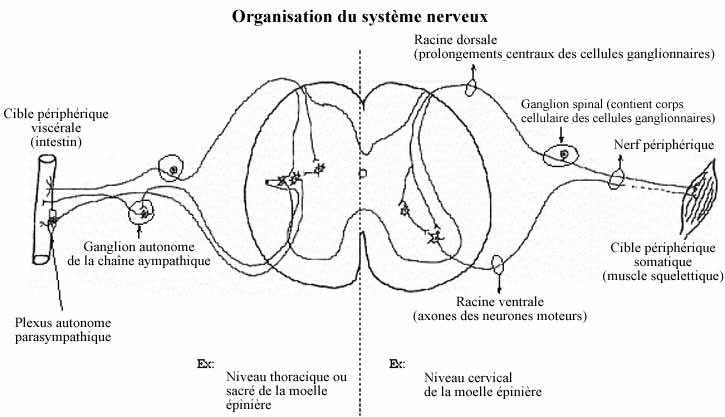
Le tissu nerveux se caractérise fonctionnellement par sa capacité de transmettre très rapidement des messages sous la forme d'une activité électrique appelée influx nerveux couplée à une activité chimique, d'une partie à l'autre de l'organisme et parfois sur de très longues distances. L'unité structurale immédiatement responsable de cette propriété est la cellule nerveuse ou neurone. Les neurones ne peuvent cependant fonctionner sans le concours des cellules gliales, qui leur procurent support et protection. Ensemble, neurones et glies permettent la production et la conduction de l'influx le long de la membrane neuronale ou neurolemme. La combinaison de plusieurs neurones dans les circuits complexes du système nerveux permet à l'animal de recevoir de l'information (stimulus) quant à son état interne, par exemple l'étirement de sa paroi gastrique, et son environnement externe, par exemple la musique ou la température, d'analyser cette information, de l'emmaganiser (mémoire), de l'intégrer avec les autres informations reçues simultanément ou antérieurement, et de répondre de manière appropriée.Voir schéma sur l'organisation du système nerveux
8.1 ORIGINE EMBRYONNAIRE
Provenant d'une portion spécifiée de l'ectoderme embryonnaire (neurectoderme), le tube neural donne naissance au système nerveux central (SNC: cerveau et moelle épinière); épithélial d'origine et quoique grandement modifié, il retiendra des caractères épithéliaux. Dorsolatérales au tube neural, les cellules de la crête neurale se détachant du neurectoderme migrent dans le mésenchyme et forment une bonne portion du système nerveux périphérique (SNP: ganglions sensoriels, ganglions et plexi autonomes du système viscéral, médulla de la surrénale, etc.).
Tant dans le tube neural (SNC) que dans la crête neurale (SNP), les cellules prolifèrent sur l'une ou l'autre lignée: neuroblastes: futurs neurones ou glioblastes: futures cellules gliales. Les neurones perdent le pouvoir de se diviser alors que les cellules gliales le conservent au besoin.
8.2 CELLULES NERVEUSES: NEURONES
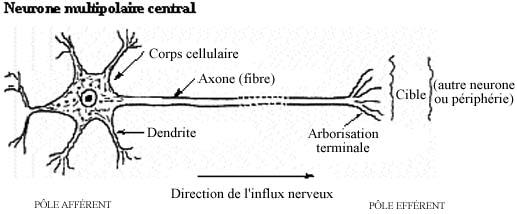
Le neurone typique est une cellule de forme multipolaire; il comprend un corps cellulaire avec noyau (caryon) et cytoplasme (péricaryon) dont émanent plusieurs prolongements cytoplasmiques qui lui confèrent la forme multipolaire. La forme (multipolaire) des neurones varie considérablement selon les types de neurones; il en est de même pour le diamètre du corps cellulaire, qui peut être aussi modeste que 6-7µm ou aussi gigantesque que 120µm. La coloration au H&E n'est guère usitée pour le tissu nerveux. Le violet de crésyl ou le bleu de toluidine, des colorants basiques, s'avèrent précieux pour décrire les corps cellulaires mais laissent les prolongements cytoplasmiques incolores. La technique d'imprégnation à l'argent tire profit de la propriété argentaphile (argyrophile) des neurones et dévoile les prolongements mais obscurcit les corps cellulaires.
8.2.1 CORPS CELLULAIRE
Le noyau (caryon) occupe généralement une position centrale dans le corps cellulaire et se caractérise par sa pâleur, mais son nucléole est gros et foncé. Les colorations dites de Nissl, au violet de crésyl ou bleu de toluidine, révèlent dans le péricaryon la présence de granules basophiles, les corps de Nissl, indiquant une grande quantité de RNA et dénotant une synthèse protéique active. En effet, les neurones contiennent plus de RNA que tout autre cellule, sauf peut-être les cellules pancréatiques. Les protéines synthétisées par les neurones sont grandement des protéines incorporées aux membranes; la conduction électrique et chimique par exocytose (voir 8.6.2) nécessitent un renouvellement membranaire considérable. Ces protéines sont transportées du corps cellulaire vers les prolongements. Un pigment jaunâtre apparaît dans les vieux neurones, les granules de lipofuchsine, dérivés de lysosomes.
8.2.2 POLARITÉ NEURONALE
Malgré ce qui a été dit sur la forme multipolaire du neurone, le terme est mal approprié et devrait être remplacé par "étoilée". Le neurone est morphologiquement et fonctionnellement polarisé et, dans ce sens, il est bipolaire. A un pôle du neurone un prolongement cytoplasmique unique, l'axone, en est le pôle émetteur; à l'autre, des prolongements multiples, les dendrites, constituent le pôle récepteur. Ce sont en réalité les dendrites qui confèrent au neurone sa forme multipolaire.
8.2.2.1 Pôle récepteur: les dendrites
Les dendrites d'un neurone sont généralement nombreuses et plus courtes que l'axone. Elles établissent des contacts fonctionnels avec d'autres neurones, en tant que récipiendaires, et constituent donc le pôle récepteur ou afférent du neurone car elles conduisent l'influx nerveux en direction centripète (par rapport au corps cellulaire).
Les dendrites peuvent s'embrancher, formant l'arborisation dendritique. Elles peuvent recevoir des contacts nombreux et de provenance multiple. Elles renferment peu de corps de Nissl et seule la portion proximale en est visible avec les colorations de Nissl. Imprégnées à l'argent, leurs parties distales apparaissent fréquemment couvertes de petites protubérances appelées épines, de minuscules évaginations cytoplasmiques servant de sites de contact fonctionnel avec les terminaisons axoniques reçues.
8.2.2.2 Pôle effecteur: l’axone
L'axone est unique à son origine et généralement assez long. A cause de sa longueur et de sa minceur relative, on nomme l'axone fibre nerveuse. Il ne faut toutefois pas la confondre avec les autres types de fibres étudiées auparavant. L'axone établit des contacts fonctionnels soit avec d'autres neurones, soit avec les organes effecteurs périphériques, i.e., les cellules musculaires, myoépithéliales et myoïdes). Il constitue le pôle effecteur ou efférent du neurone car il conduit l'influx de façon centrifuge (par rapport au corps cellulaire) et contrôle l'activité de ses cibles.
Le neurone dont l'axone quitte le SNC est un neurone moteur (motoneurone); s'il innerve directement un muscle on l'appelle motoneurone somatique, car il s'agit d'un muscle strié squelettique (revoir 7.1), mais s'il innerve un ganglion autonome qui, lui, innerve les muscles cardiaque et lisse, on l'appelle motoneurone viscéral. Le neurone dont l'axone pénètre dans le SNC (afférence primaire) est un neurone sensoriel et son corps cellulaire se retrouve dans un ganglion sensoriel à l'extérieur du SNC. Le neurone du SNC dont l'axone innerve d'autres neurones du SNC demeure, de ce fait, confiné au SNC. Ce sont les plus nombreux. Parmi les neurones centraux, on distingue les neurones de projection, dont l'axone se projette loin du corps cellulaire, et les interneurones locaux, dont le court axone innerve des cibles à proximité du corps cellulaire d'origine.
L'axone est issu d'une région spécialisée du corps cellulaire ou d'une dendrite primaire, le segment initial. Bien qu'unique à son origine, l'axone peut se subdiviser sur son parcours et projeter ainsi des collatérales vers différentes cibles. Selon le type de neurone, sa longueur peut atteindre quelques dizaines de micromètres ou plusieurs mètres. A l'abord de sa cible, chaque collatérale se ramifie, formant l'arborisation terminale (autrefois appelée télodendrie). L'axone ne contenant pas de corps de Nissl, il demeure invisible avec les colorations de Nissl.
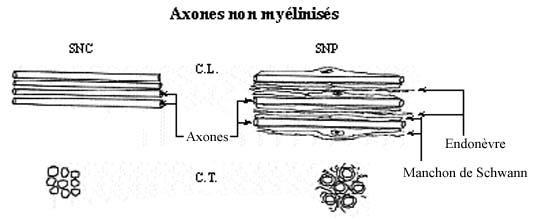
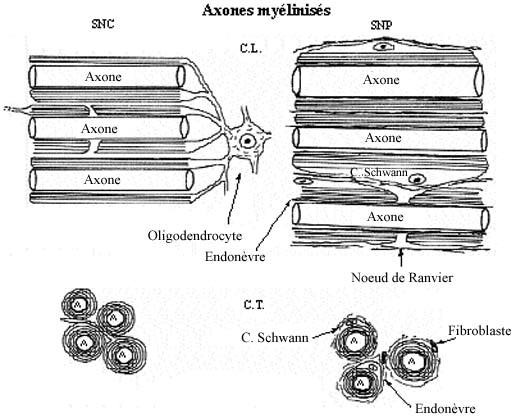
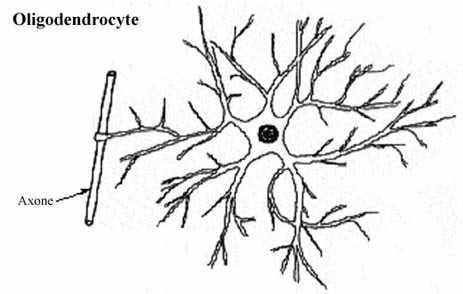
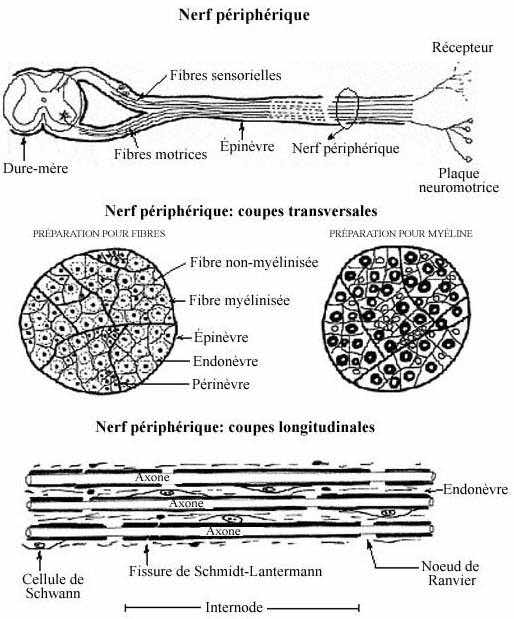
Il existe toute une gamme de diamètre axonique. Les axones de moins de 1µm de diamètre sont généralement non myélinisés, ceux plus gros le sont. La gaine de myéline entourant les axones est fabriquée par des cellules gliales, les oligodendrocytes dans le SNC et les cellules de Schwann dans le SNP, de la façon suivante. La cellule gliale étend un prolongement cytoplasmique qu'elle enroule maintes fois autour de l'axone et, ce faisant, retire le cytoplasme du prolongement, n'y laissant qu'une enveloppe membranaire plusieurs fois enroulée. Cette membrane cellulaire qui forme la myéline a une composition à forte teneur lipidique.
Le manchon de myéline est interrompu à intervalles réguliers le long de l'axone, les noeuds de Ranvier, qui ressemblent à des étranglements en microscopie photonique. La portion myélinisée entre deux noeuds est le segment internodal. La myéline isole électriquement l'axone; en empêchant la transmission d'influx nerveux, celui-ci doit "sauter" d'un noeud de Ranvier à l'autre. Ce phénomène de la conduction saltatoire accélère énormément la conduction électrique le long de l'axone. Notons que les dendrites sont dépourvues de myéline.
Voir schéma sur les axones non myélinisés et les axones myélinisés
8.3 CELLULES GLIALES
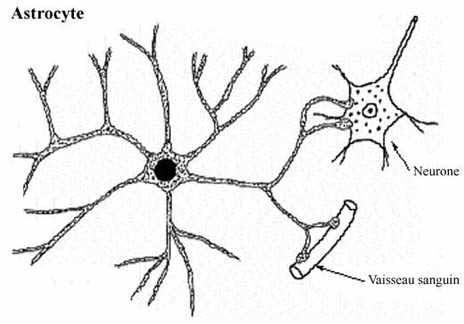
Beaucoup plus nombreuses que les neurones, les cellules gliales ne se laissent pas facilement étudier. Au Nissl, seul leur noyau ressort, comme c'est le cas des fibroblastes du tissu conjonctif. Des imprégnations métalliques mettent en évidence leurs corps cellulaires et leurs nombreux bras cytoplasmiques. Il devient possible d'apprécier leurs relations topographiques avec les neurones et les vaisseaux sanguins.
La microscopie électronique dévoile que l'espace intercellulaire est réduit au minimum dans le SNC. Les cellules gliales remplissent l'espace entre les neurones (incluant leurs prolongements cytoplasmiques) et les vaisseaux sanguins, et s'interposent derechef entre neurones et pie-mère, la méninge la plus interne. En empêchant tout attouchement entre membrane neuronale et tout tissu non neural, elles isolent physiquement les cellules nerveuses. Toute substance qui gagne le neurone doit d'abord traverser la barrière gliale. Les cellules gliales assurent l'homéostasie du milieu dans lequel vivent les neurones; elles tamponnent le milieu s'il se crée un déséquilibre ionique. Ceci revêt une importance capitale pour la fonction hautement spécialisée qu'accomplissent les neurones.
Selon la terminologie classique, il existe deux grandes classes de cellules gliales: macroglie et microglie. Les cellules microgliales se dissocient des autres tant en ce qui concerne leur morphologie que leur fonction ou leur origine. Elles proviendraient en effet des monocytes du sang. Peu nombreuses en situation normale, elles prolifèrent lors d'endommagement au SNC pour jouer le rôle de macrophage.
8.3.1 CELLULES GLIALES DU SNC
Les cellules macrogliales du SNC dérivent des glioblastes du tube neural embryonnaire. Elles conservent la propriété de se diviser chez l'animal adulte, surtout lors de trauma, afin de protéger les neurones des envahisseurs étrangers.
Les plus grosses et plus nombreuses des cellules gliales, les astrocytes possèdent des bras cytoplasmiques qui leur confèrent une apparence étoilée. Ce sont surtout les astrocytes qui assument les rôles de tampon du milieu et de protecteur des neurones.
Plus petits et moins nombreux que les astrocytes, les oligodendrocytes se caractérisent particulièrement par leur fonction de myélinisation des axones du SNC (revoir 8.2.2.2).
8.3.2 CELLULES GLIALES DU SNP
Les cellules gliales du SNP prennent origine des glioblastes de la crête neurale de l'embryon. Elles aussi conservent la propriété de se diviser.
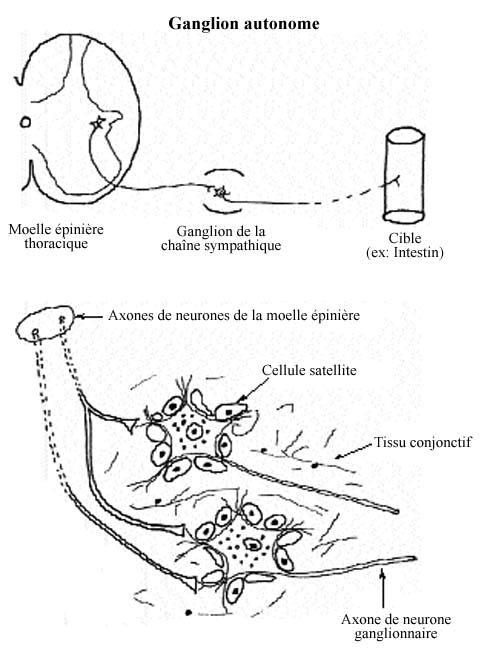
Les cellules satellites qui entourent les neurones ganglionnaires, tant sensoriels qu'autonomes, remplissent une fonction analogue à celle des astrocytes, mais dans un milieu où le tissu non-neural abonde.
Analogues aux oligodendrocytes, les cellules de Schwann myélinisent les fibres nerveuses périphériques. Plusieurs différences existent entre les deux types cellulaires de même qu'entre les deux types de gaine de myéline produite, mais ces détails ne seront pas exposés dans ce cours.
8.4 SYSTEME NERVEUX CENTRAL (SNC)
Le système nerveux central comprend le cerveau et la moelle épinière qui forment ensemble le névraxe. Le névraxe est enveloppé de trois couches conjonctives, les méninges. La plus interne et la plus délicate, la pie-mère, s'insinue dans le SNC et recouvre jusqu'aux plus petits vaisseaux sanguins irriguant le tissu. L'arachnoïde, méninge moyenne, renferme de nombreux vaisseaux sanguins. Elle s'entoure d'une solide capsule conjonctive, la dure-mère.
Une coupe fraîche du SNC révèle deux zones principales, matière grise et matière blanche, également distinctes sur coupe histologique. La matière blanche est faite d'axones qui se regroupent en faisceaux, ou voies nerveuses, pour atteindre des cibles parfois lointaines du névraxe. La couleur blanche incombe à la myéline autour de nombreux axones, riche en lipides membranaires. La matière grise est grise par manque de myéline. Elle regroupe en effet les corps cellulaires de neurones qui ont tendance à s'agréger en "noyaux" ou en couches.
Dans la moelle épinière, la matière grise occupe une position centrale autour du canal épendymaire et adopte la forme d'un papillon, tel que vu en coupe transversale, entouré par la matière blanche. Dans le cerveau, les corps cellulaires se regroupent en agrégats, ou noyaux (ne pas confondre avec noyau -caryon- cellulaire), séparés les uns des autres par de la matière blanche, ou en couches, parfois en alternance avec des couches de matière blanche.
En relation avec leur fonction respective, les astrocytes se rencontrent davantage dans la matière grise, mais leurs prolongements cytoplasmiques s'étendent dans la matière blanche, et les oligodendrocytes se retrouvent davantage dans la matière blanche.
La majorité des neurones du SNC sont des neurones intercalés entre les afférences primaires et les neurones moteurs. Les afférences primaires établissent parfois des contacts directement avec les dendrites de motoneurones, mais établissent plus souvent des contacts avec ces neurones centraux. En fait, plusieurs neurones centraux peuvent être intercalés entre les afférences primaires et les motoneurones. De formes multipolaires variées et plus petits que les motoneurones, les neurones centraux sont donc confinés au SNC.
8.5 SYSTEME NERVEUX PERIPHERIQUE
8.5.1 GANGLIONS SENSORIELS: CRANIENS ET SPINAUX (rachidiens)
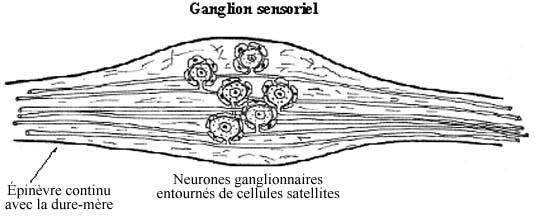
Les ganglions sensoriels incluent les ganglions spinaux situés latéralement à la moelle épinière et les ganglions crâniens à des positions crâniennes à peu près correspondantes. Ils renferment les corps cellulaires de neurones sensoriels, les seuls qui reçoivent directement les stimuli (goût, douleur cutanée ou viscérale, etc., etc.). Ces neurones ganglionnaires sensoriels traduisent les stimuli en influx nerveux qu'ils acheminent vers le SNC. Pour ce faire, le neurone ganglionnaire sensoriel possède une organisation morphologique bien particulière.
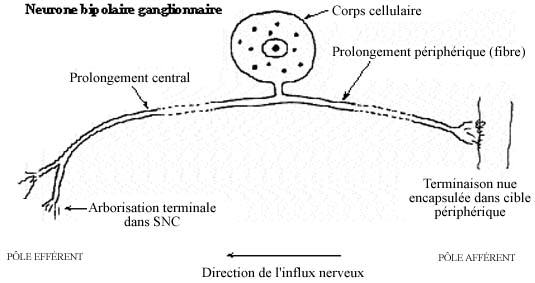
Il est de forme bipolaire, n'ayant que deux prolongements cytoplasmiques qui prennent naissance d'une même région du corps cellulaire, donnant l'impression d'un prolongement unique qui bifurque en T (pseudo-unipolaire). L'un d'eux tient lieu de dendrite et constitue le pôle afférent de la cellule. Sans s'embrancher comme une dendrite ordinaire, il s'étend jusqu'à la périphérie et sa terminaison, qui peut s'arboriser, prend le nom de récepteur (voir 8.5.2). C'est ce dernier qui perçoit le stimulus. Le prolongement périphérique, même si analogue à une dendrite, ne devrait pas en porter le nom. Il est d'ailleurs très long et mérite d'emblée l'épithète de fibre nerveuse. De surcroît, il peut être myélinisé. Les prolongements périphériques provenant d'un même ganglion se réunissent en un faisceau compact qui se subdivise distalement à l'abord de leurs cibles diverses.
Le prolongement central, diamétralement opposé au premier, se projette vers le SNC et constitue l'afférence primaire. Analogue à l'axone, c'est le pôle effecteur de la cellule sensorielle car il agit sur les neurones du SNC. A l'instar du prolongement périphérique, il peut être myélinisé. Les prolongements centraux d'un ganglion spinal constituent ensemble la racine dorsale pénétrant dans la corne dorsale de la moelle épinière.
Les ganglions sensoriels renferment les cellules satellites, petites et aplaties, encerclant le corps cellulaire de chaque cellule nerveuse. On retrouve des vaisseaux sanguins et, en contraste avec le SNC, beaucoup de tissu conjonctif.
8.5.2 RECEPTEURS
C'est donc le prolongement périphérique du neurone ganglionnaire sensoriel qui reçoit l'information. Les stimuli appartiennent à une variété de modalités: goût, étirement musculaire et tendineux, douleur somatique (musculaire, cutanée) et viscérale, toucher, etc. Le récepteur agit comme traducteur de l'information périphérique, quelle qu'elle soit, en influx nerveux, seul langage compris par les neurones. Sa configuration, parfois complexe, est reliée à la modalité sensorielle. Il existe plusieurs classifications des récepteurs: selon leur distribution, leur morphologie, leur modalité sensorielle, la rapidité de conduction de leur fibre, etc. Nous adopterons ici une classification morphologique très simple, qui scinde les récepteurs en deux grands groupes.
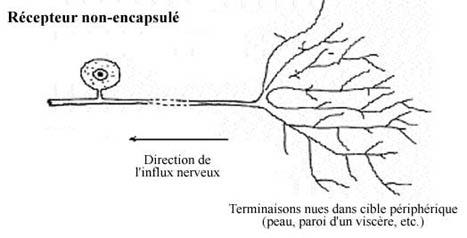
8.5.2.1 Récepteurs non-encapsulés ou terminaisons nues
Chaque terminaison de l'arborisation terminale d'un prolongement périphérique est "nue": le contact avec le stimulus est direct. Les plus communes, elles se retrouvent dans toutes les parties de l'organisme. Leur vitesse de conduction lente et leur champ récepteur étendu font qu'elles transmettent des sensations dites diffuses, par exemple la douleur, la température. La terminaison nerveuse associée au follicule pileux (voir 9.4) entre dans cette catégorie.
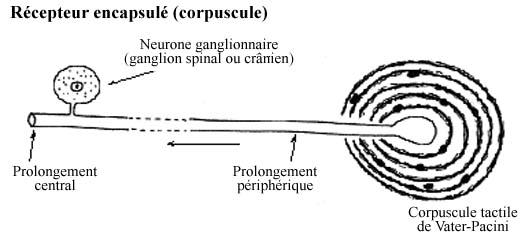
8.5.2.2 Récepteurs encapsulés: Corpuscules
Les récepteurs encapsulés présentent une certaine complexité morphologique et des variations. La terminaison du prolongement périphérique s'associe à des cellules conjonctives modifiées. La terminaison nerveuse et sa capsule, ensemble, forment le récepteur: le contact avec le stimulus s'effectue par la capsule. Ce type de récepteurs a été traditionnellement nommé corpuscule. Leur vitesse de conduction plus rapide et leur champ récepteur restreint font que les corpuscules transmettent des sensations précises, discriminantes, telles le toucher de précision. Entrent dans cette catégorie les corpuscules de Krause de la conjonctive de l'oeil, les corpuscules de Meissner et de Vater-Pacini de la peau, les fuseaux neuromusculaires des muscles squelettiques, les organes neurotendineux de Golgi.
Les récepteurs des organes sensoriels spéciaux (olfaction, vision, audition, équilibre) comprennent, en plus des prolongements périphériques de neurones ganglionnaires, un épithélium spécialisé. Ce sont les cellules épithéliales qui sont stimulées et leur excitation est transmise aux terminaisons nerveuses. Ces récepteurs fort complexes ne seront pas détaillés ici.
8.5.3 NERF PERIPHERIQUE (voir schéma)
A la sortie de la moelle épinière, à chaque segment, les axones des motoneurones forment la racine ventrale qui jouxte ventralement le ganglion sensoriel et ses prolongements périphériques. Ainsi, les fibres motrices de la racine ventrale et les fibres sensorielles des ganglions parcourent ensemble leur route périphérique; elles forment ainsi le nerf périphérique. La situation est semblable en ce qui concerne les nerfs crâniens sauf que tous ne sont pas mixtes: certains d'entre eux sont exclusivement sensoriels, d'autres exclusivement moteurs. Le nerf se subdivise à maintes reprises à mesure que des groupes de fibres atteignent leur cible respective. Le nerf s'accompagne souvent des vaisseaux sanguins et lymphatiques qui desservent les mêmes territoires.
Le nerf entier s'entoure d'un dense manchon de tissu conjonctif collagénique et élastique, l'épinèvre. Du tissu conjonctif moins dense et continu avec le précédent, le périnèvre, subdivise le nerf. Chaque fibre nerveuse est elle-même entourée de délicat tissu conjonctif, continu avec le précédent, l'endonèvre. Ces enveloppes conjonctives recouvrent aussi le ganglion sensoriel et les racines dorsale et ventrale, et elles sont continues avec les trois méninges protégeant le SNC.
Il faut comprendre la différence entre une collection d'axones centraux, appelée faisceau ou voie, et celle de fibres périphériques, appelée nerf. Le nerf renferme du tissu conjonctif, le faisceau central non. Le nerf comprend des fibres dites primaires (motrices et sensorielles) tandis que la plupart des voies centrales (pas toutes) comprennent des fibres (axones) de neurones centraux.
Les fibres motrices et sensorielles du nerf peuvent être myélinisées. La myéline consistant grandement en lipides membranaires, il va de soi que les préparations ordinaires la dissoudent. Voilà pourquoi il faut déterminer quelles composantes nous intéressent avant d'entreprendre l'étude du nerf périphérique, de façon à choisir le procédé adéquat. Pour préserver la myéline, il faut éviter d'employer l'éthanol, l'éther ou le chloroforme. La myéline intacte se colore au bleu Luxol et apparaît comme un anneau bleu ayant un centre incolore. En effet, la fibre se colore rarement. Les colorations utilisant le nitrate d'argent imprègnent les fibres mais pas la myéline.
8.5.4 GANGLIONS ET PLEXI DU SYSTEME NERVEUX VISCÉRAL
Nous avons peu insisté sur l'existence de deux composantes fonctionnelles du système nerveux, soit la composante somatique et celle viscérale. La première contrôle la musculature squelettique et est dite volontaire. La seconde contrôle la musculature lisse et cardiaque et est dite involontaire. Alors que les motoneurones somatiques projettent directement leurs axones à leurs cibles musculaires, les motoneurones viscéraux envoient leurs axones à l'un ou l'autre relai intermédiaire: soit un ganglion autonome, soit un plexus. Chacun des deux types de relai comprend les mêmes composantes anatomiques et fonctionnelles mais les ganglions composent des structures bien distinctes alors que les plexi sont des structures diffuses tout près de la paroi des organes cibles ou même dedans la paroi.
Le neurone ganglionnaire autonome est formé d'un corps cellulaire de forme multipolaire (étoilée) à partir duquel émanent des dendrites et un axone. Les dendrites reçoivent les terminaisons axoniques des motoneurones viscéraux (revoir 8.2.2.2). L'axone quitte le ganglion ou le plexus pour innerver ses cibles: cellules musculaires lisses ou cardiaques, myoépithéliales et myoïdes. Les neurones ganglionnaires bénéficient de l'entourage des cellules satellites.
Ainsi, les ganglions et plexi autonomes diffèrent des ganglions sensoriels en étant le site de contacts synaptiques entre axones et dendrites, et ils renferment des interneurones. Leurs neurones ne sont pas sensoriels.
8.6 CONTACTS FONCTIONNELS DES NEURONES
Les neurones sensoriels traduisent les signaux périphériques en influx nerveux qu'ils conduisent au SNC où, grâce à des contacts morphologiques et fonctionnels, ils communiquent avec les neurones centraux. Plusieurs neurones centraux établissent des contacts entre eux, en rapport avec l'information périphérique reçue. Ces échanges forment la base de l'analyse et l'intégration de l'information. Eventuellement, plusieurs interneurones convergent vers les motoneurones et leur transmettent un message intégré, pour que le travail de ces derniers sur les organes effecteurs constitue une réponse appropriée. Le dernier niveau du transfert d'information s'effectue donc entre l'axone somato- ou viscéromoteur et la cible périphérique. Tout au long de cet arc, le passage de l'influx d'un neurone à l'autre puis à la cellule musculaire, se fait au niveau des contacts morphologiques et fonctionnels précités, contacts très spécialisés connus sous le nom de synapses. Il existe deux types fonctionnels de synapse: chimique et électrique.
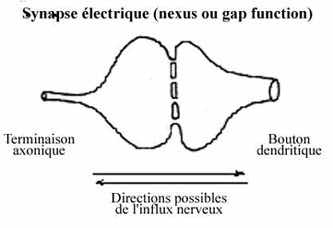
8.6.1 "SYNAPSE" ELECTRIQUE OU NEXUS
La "synapse" électrique est ni plus ni moins une jonction cellulaire du type nexus ("gap junction"); elle s'opère par continuité membranaire entre deux neurones, par le biais de petits tunnels. La vague de dépolarisation de la membrane se transmet directement à la membrane de l'autre neurone. Ce contact est bidirectionnel et peu sélectif. Le terme synapse ne leur sied pas parfaitement.
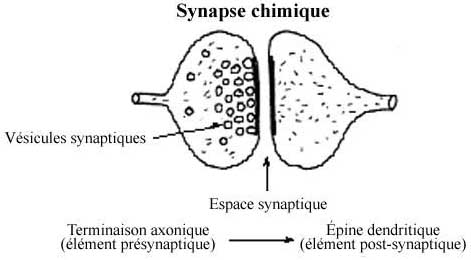
8.6.2 SYNAPSE CHIMIQUE
La synapse chimique s'accomplit à un site de jonction cellulaire de type adherens modifié, entre la terminaison axonique d'un neurone: l'élément présynaptique, et la dendrite d'un autre neurone: l'élément postsynaptique. Il n'y a pas continuité membranaire entre les deux éléments, qui ménagent un espace minime entre eux au site jonctionnel, la fente synaptique.
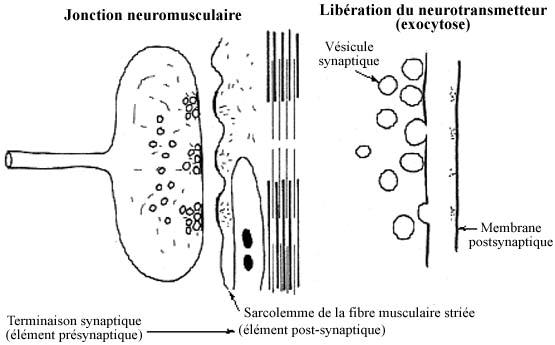
Une vague de dépolarisation (l'influx nerveux) le long de la membrane présynaptique déclenche la libération par la terminaison d'un produit chimique, le neurotransmetteur, dans la fente synaptique. Le neurotransmetteur est en réserve dans des petites vésicules axoplasmiques, les vésicules synaptiques, qui, sous l'effet de la vague de dépolarisation, fusionnent avec la membrane présynaptique et le libèrent par exocytose. Une fois dans la fente synaptique, le neurotransmetteur peut se lier à des récepteurs de la membrane postsynaptique et altérer le potentiel membranaire du neurone postsynaptique. Certains neurotransmetteurs ont un effet excitateur, d'autres un effet inhibiteur sur l'élément postsynaptique.
Comme un neurone donné reçoit toute une batterie d'afférences, tant excitatrices qu'inhibitrices, en proportions diverses et sur des sites dendritiques différents, c'est la sommation de toutes les afférences à un instant donné qui détermine si ce neurone produira à son tour une vague de dépolarisation (potentiel d'action) ou sera plutôt inhibé.
Quelques neurotransmetteurs bien connus: acétylcholine, dopamine, norépinéphrine (noradrénaline), sérotonine, GABA (gamma amino butiric acid), glutamate, substance P.
Les techniques modernes d'immunohistochimie permettent de démontrer la présence d'un neurotransmetteur, ou de son précurseur biochimique, à l'intérieur de neurones. D'autre part, les techniques de "binding" s'utilisent pour déceler sur les membranes neuronales la présence de récepteurs pour des neurotransmetteurs donnés. Nous arrivons à une cartographie biochimique du SNC et du SNP de plus en plus raffinée.
Le contact qu'établit l'axone avec la cellule musculaire est de type chimique et ressemble morphologiquement à la synapse chimique juste décrite. On l'appelle jonction neuromusculaire ou plaque neuromotrice. Elle est toujours excitatrice et utilise le neurotransmetteur acétylcholine.
Les synapses ne se voient pas en microscopie photonique. Toutefois, à cause de la rareté des attouchements entre neurones, il est tentant de déduire que lorsqu'une terminaison axonique jouxte une épine dendritique, comme on peut le voir dans certaines préparations à l'argent, cela suggère fortement l'occurrence d'une synapse. L'examen d'une telle figure en microscopie électronique confirme le cas.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}