FÉCONDATION ou FERTILISATION (cours 3)
La fécondation, ou fertilisation, est le processus selon lequel les gamètes mâle et femelle, spermatozoïde et oeuf, fusionnent pour créer un nouvel individu, le zygote, avec des potentiels génétiques dérivés des deux parents.
La fécondation accomplit donc deux activités séparées:
le sexe: combinaison des gènes de deux individus pour les transmettre au rejeton, et
la reproduction: création d'un nouvel organisme.
La fertilisation comprend 4 étapes majeures:
1- reconnaissance et contact du spermatozoïde et de l'oeuf, étape du contrôle de la qualité. Les deux gamètes doivent appartenir à la même espèce.
2- entrée du spermatozoïde dans l'oeuf, étape du contrôle de la quantité. Un seul spermatozoïde peut fertiliser l'oeuf, tous les autres devant être éliminés.
3- amphimixie, fusion des noyaux des deux gamètes, donc de leur matériel génétique.
4- activation du métabolisme de l'oeuf pour entamer le développement embryonnaire.
Rôle du spermatozoïde dans la fertilisation?
Nous prenons maintenant pour acquis le fait que le spermatozoïde est essentiel à la fécondation de l'oeuf, mais il n'en a pas toujours été ainsi. Ce n'est qu'au cours des 100 dernières années que son rôle dans la fécondation a été élucidé.
- 17e siècle:
Anton van Leuwenhoek, le microscopiste hollandais co-découvreur des spermatozoïdes en 1678, les croyait des animaux parasites du sémen, d'où leur nom: animaux du sperme. On les appelait également animalcules. Selon Leuwenhoek ils n'avaient rien à voir avec la reproduction. Nicolas Hartsoeker, l'autre co-découvreur, soumit une hypothèse alternative: l'embryon était préformé dans la tête du spermatozoïde, hypothèse qui gagna alors peu de popularité; les scientifiques ont appuyé van Leuwenhoek.
-18e siècle:
Vers la fin du 18e siècle, Lazarro Spallanzani démontra que du sperme filtré, démuni de spermatozoïdes, ne fécondait pas l'oeuf de crapaud. Il conclut néanmoins que le sémen était l'agent fertilisateur, les spermatozoïdes n'étant que des animaux parasites!
- 19e siècle:
En 1821, J.B. Dumas et J.L. Prévost affirmèrent que les spermatozoïdes étaient les agents actifs de la fertilisation à cause:
de leur existence universelle chez les mâles sexuellement mûrs,
de leur absence chez les individus immatures et âgés, et
de leur absence chez le mulet stérile.
Cette déclaration fut néanmoins ignorée jusque vers 1840 alors que A. von Kolliker décrivit la formation des spermatozoïdes à partir de cellules testiculaires et conclut qu'ils étaient des cellules grandement modifiées. Il nia toutefois qu'il existe un contact entre le spermatozoïde et l'oeuf lors de la fécondation; le spermatozoïde ne fait qu'exciter l'oeuf.
En 1876 Oscar Hertwig décrivit la pénétration de l'oeuf par le spermatozoïde et l'union de leurs noyaux. Il nota qu'un seul spermatozoïde pénètre l'oeuf et que les cellules de l'embryon dérivent toutes des deux cellules fusionnées à la fertilisation.
Herman Fol fournit par la suite plus de détails sur le mécanisme de la pénétration de l'oeuf d'oursin par le spermatozoïde.
Etapes de la fécondation:
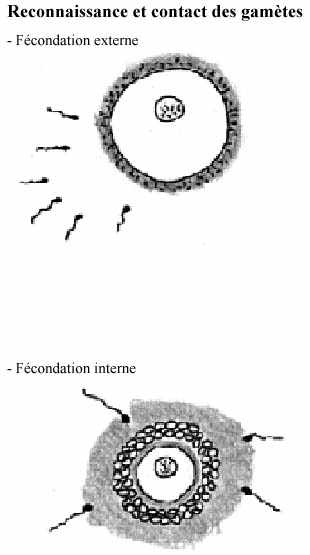
1- Reconnaissance et contact du spermatozoïde et de l'oeuf:
Plusieurs animaux aquatiques et amphibiens libèrent leurs gamètes dans l'environnement local; ils n'ont pas recours à la fécondation interne. Il existe des mécanismes qui permettent aux spermatozoïdes de retrouver les oeufs et qui empêchent la fécondation interspécifique. Des substances chimiques libérées par la gelée entourant l'oeuf de ces animaux agissent à distance, par chimiotaxie, en augmentant la motilité des spermatozoïdes et les attirant vers l'oeuf.
Des glycoprotéines membranaires de la gelée appelées fertilisines induisent l'agglutination des spermatozoïdes à la surface de l'oeuf. Les spermatozoïdes s'y attachent grâce à des récepteurs de surface appelés antifertilisines. La réaction fertilisine - antifertilisine est relativement spécifique; elle peut se produire entre espèces voisines mais est faible ou nulle entre espèces éloignées. La gelée glycoprotéique qui entoure les oeufs des animaux aquatiques se dissout progressivement dans l'eau; des oeufs longtemps laissés dans l'eau deviennent non fécondables.
La fécondation externe ne relève pas complètement du hasard. Des rapprochements, presque des accouplements, entre mâle et femelle peuvent avoir lieu; la maturation sexuelle et l'émission des gamètes peuvent être simultanées et dépendent alors des saisons, des cycles lunaires ou autres; la ponte peut déclencher l'émission des spermatozoïdes.
Les animaux terrestres pratiquent la fécondation interne. L'ovulation est contemporaine ou proche de l'insémination des spermatozoïdes; cette simultanéité est d'autant plus importante que la durée de vie des gamètes émises est courte: quelques heures à quelques jours. Les voies génitales femelles jouent un rôle actif dans la fertilisation; leur milieu acide, normalement défavorable à la survie des spermatozoïdes, devient plus alcalin avant l'ovulation grâce à des sécrétions muqueuses utérines. Des centaines de milliers ou millions de spermatozoïdes émis, seulement quelques milliers franchissent l'utérus et ce juste avant l'ovulation. Leur progression vers les oviductes s'effectue par le mouvement des flagelles, aidé de mouvements de l'oviducte. Quelques centaines de spermatozoïdes seulement arriveront à l'ovule. Il n'est pas certain que la chimiotaxie intervienne dans l'attraction du spermatozoïde vers l'oeuf. La gelée glycoprotéique libérée à l'ovulation capte les spermatozoïdes.
Le spermatozoïde mammalien est incapable de fertiliser l'oeuf s'il n'a pas préalablement résidé un certain temps dans les voies femelles, où il subit la capacitation. Celle-ci consiste en changements de la structure lipidique de sa membrane cellulaire, dont du taux cholestérol / phospholipides qui décroît. Les molécules d'albumine retrouvées dans les voies femelles enlèvent du cholestérol à la membrane du spermatozoïde, ce qui la déstabilise.
2- Entrée du spermatozoïde dans l'oeuf:
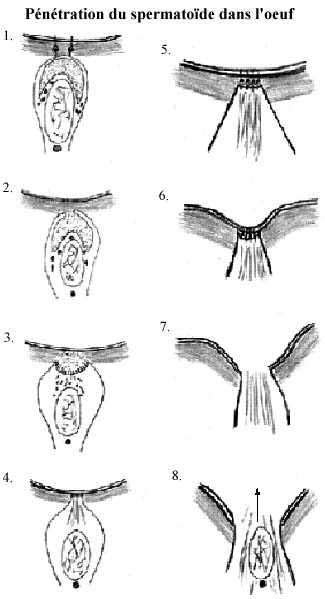
Chez les animaux aquatiques, le contact du spermatozoïde avec la gelée entourant l'oeuf initie un processus appelé réaction acrosomienne, à deux composantes: la rupture de l'acrosome et l'émission du prolongement acrosomien.
Un polysaccharide sulfaté de la gelée de l'oeuf affecte la membrane de la tête du spermatozoïde et permet l'entrée de Ca++ et Na+ et la sortie de K+ et H+. Ce transfert ionique amène la fusion de la membrane cytoplasmique de la tête avec la membrane de l'acrosome sous-jacent, puis la rupture des membranes fusionnées. Les enzymes protéolytiques de l'acrosome sont ainsi libérées et peuvent digérer la gelée de l'oeuf.
Le prolongement acrosomien prend naissance de la polymérisation des molécules globulaires d'actine en filaments d'actine, dépendante de la libération d'ions H+ par la tête du spermatozoïde. On l'appelle aussi tubule ou filament acrosomien. Le prolongement acrosomien entre en contact avec l'enveloppe vitelline de l'oeuf, contact médié par une protéine acrosomienne appelée bindine et pour laquelle se trouve un récepteur sur l'enveloppe vitelline. La réaction bindine et son récepteur est hautement spécifique.
Le contact du tubule acrosomien avec l'enveloppe vitelline produit une réaction locale de la membrane de l'oeuf qui se soulève pour former le cône de fertilisation. La membrane du cône se rompt et fusionne avec celle du spermatozoïde (déjà rompue). Le cytoplasme de l'oeuf s'insinue dans le spermatozoïde et englobe son noyau et le centriole de la pièce intermédiaire, propulsés par la contraction du prolongement acrosomien. Le reste du spermatozoïde se détache et dégénère.
Pour les animaux à fécondation interne, on ne parle pas de réaction acrosomienne. Chez certains, la déstabilisation membranaire subie lors de la capacitation dans les voies femelles cause la rupture de l'acrosome et la libération des enzymes quand le spermatozoïde est encore à une certaine distance de l'ovule. Les enzymes brisent les liens intercellulaires qui cimentent les cellules folliculaires, permettant aux spermatozoïdes d'atteindre l'enveloppe vitelline (ou la zone pellucide) autour de l'ovocyte. Chez d'autres, les spermatozoïdes arrivent à traverser la corona radiata avant le bris de l'acrosome, qui ne se produit qu'une fois le spermatozoïde attaché à l'enveloppe vitelline.
Une étape de la capacitation consiste en démasquage d'une enzyme de surface, la glycosyltransférase, qui reconnaît un carbohydrate N-acétylhexosamine sur la zone pellucide. La spécificité de l'attachement du spermatozoïde à la zone pellucide n'est pas très élevée, mais les risques de fécondation interspécifique chez ces animaux sont faibles.
L'acrosome contient une enzyme, l'acrosine, dont la séquence en acides aminés ressemble à celle de la trypsine et qui digère la zone pellucide au point d'attachement. Comme la trypsine, elle doit être activée pour devenir fonctionnelle, ce qui se produit par une glycoprotéine retrouvée dans les voies femelles. Il s'ensuit une fusion des membranes cytoplasmiques du spermatozoïde et de l'oeuf. Ce n'est pas nécessairement la membrane de l'apex de la tête qui fusionne, mais celle de la région latérale.
Compte tenu de la polarité de l'oeuf, particulièrement de l'hétérogénéité membranaire, il est probable que toute sa surface n'ait pas le même pouvoir de liaison avec le spermatozoïde. On sait que chez la grenouille, le spermatozoïde pénètre l'oeuf préférentiellement à l'équateur; chez la souris, la fusion de la membrane du spermatozoïde s'accomplit préférentiellement avec la région lisse de la membrane de l'oeuf (soit à l'antipode du point d'émission du premier globule polaire).
Chez l'oursin, où le phénomène a été particulièrement bien étudié, et chez des amphibiens, deux mécanismes: l'un rapide et l'autre lent, gardent contre la polyspermie.
réaction rapide:
Un spermatozoïde ne peut fusionner avec la membrane de l'oeuf que lorsqu'elle est à son potentiel de repos: -70mV. Dès qu'un spermatozoïde pénètre, une protéine de l'acrosome fait ouvrir les canaux ioniques de la membrane de l'oeuf et permet l'entrée de Na+. Or, l'entrée de Na+ dans l'oeuf, qui en contient peu comparativement au milieu marin dans lequel il baigne, élève le potentiel à 0 à +20mV. L'élévation de potentiel membranaire empêche d'autres spermatozoïdes de fusionner avec la membrane de l'oeuf.
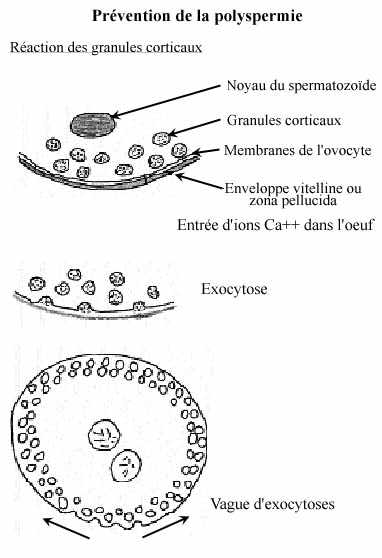
réaction lente:
Le potentiel membranaire de l'oeuf retourne au repos alors que plusieurs spermatozoïdes y sont encore accolés. Un mécanisme lent les y enlève: la réaction des granules corticaux. Elle ressemble à la réaction acrosomienne: en présence d'ions Ca++ la membrane des granules fusionne avec celle de l'ovocyte, libérant leur contenu dans l'espace entre la membrane de l'ovocyte et l'enveloppe vitelline. Suivant le bris des granules corticaux au point d'entrée du spermatozoïde, une vague d'exocytose des granules corticaux se propage tout autour de l'oeuf.
Les enzymes protéolytiques libérées dissoudent les protéines liant l'enveloppe vitelline à la membrane cellulaire, élargissant l'espace entre elles: l'espace périvitellin. L'enveloppe vitelline, distancée de la membrane cellulaire, devient l'enveloppe de fertilisation, qui se forme au site d'entrée du spermatozoïde et progresse tout autour de l'oeuf, étant complétée en quelques secondes ou plusieurs minutes, selon les espèces. L'enveloppe de fertilisation subit des changements: des protéases, libérées par les granules corticaux, inactivent ou même arrachent les récepteurs de spermatozoïdes, puis une peroxydase la fait durcir. L'enveloppe de fertilisation durcie offre ainsi une barrière mécanique.
Les ovocytes de la plupart des espèces utilisent tant le blocage rapide que le blocage lent pour prévenir la polyspermie. Chez les mammifères, la polyspermie est minimisée par le petit nombre de spermatozoïdes atteignant l'oeuf. La réaction des granules corticaux ne crée pas d'enveloppe de fertilisation mais l'effet est le même: la libération d'enzymes modifie les récepteurs spermatiques de la zone pellucide de sorte qu'ils ne lient plus les spermatozoïdes. C'est la réaction de la zona pellucida.
Chez certains oiseaux, reptiles et amphibiens, plusieurs spermatozoïdes pénètrent l'oeuf mais, par un mécanisme inconnu, tous sauf un se désintègrent après l'amphimixie.
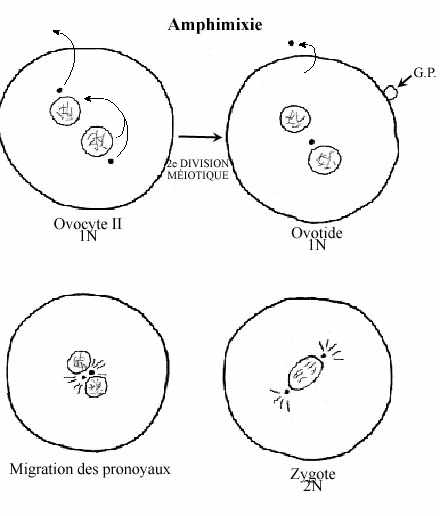
3- Amphimixie: fusion du matériel génétique et rétablissement de la diploïdie:
Après fusion des membranes de l'ovocyte et du spermatozoïde, le noyau et le centriole de ce dernier se séparent des mitochondries et du flagelle qui se désintègrent. Les mitochondries du zygote ne proviennent que de l'oeuf. Une fois dans l'oeuf, la chromatine du noyau du spermatozoïde se décondense très rapidement, la membrane nucléaire se disperse et le volume du noyau augmente; celui-ci porte le nom de pronoyau mâle. Une fois haploïde, le noyau de l'oeuf porte le nom de pronoyau femelle, lui aussi à chromatine décondensée et volume accru.
Le pronoyau et le centriole mâles subissent une rotation de 180° de sorte que le centriole mâle fait face au pronoyau femelle. Le pronoyau femelle complète sa deuxième division méiotique, avec émission du second globule polaire et expulsion du centriole. Le centriole mâle se dédouble et les deux s'éloignent du centre du cytoplasme; les microtubules centriolaires forment deux asters. Les deux pronoyaux migrent l'un vers l'autre; à leur rencontre, les membranes nucléaires se brisent, fusionnent et une membrane nucléaire unique se forme.
Chez l'oursin un véritable noyau zygote diploïde est formé. Les chromosomes paternels et maternels s'individualisent à nouveau, s'apparient et s'organisent à l'équateur du fuseau achromatique au moment de la première division de segmentation (mitotique). Chez les mammifères, par contre, la chromatine se condense en chromosomes qui s'orientent le long du fuseau mitotique commun pendant tout le stade zygote. Le mécanisme de fusion nucléaire prend environ 1 heure chez l'oursin et 12 heures chez l'humain.
Dans les oeufs alécithes et oligolécithes, les deux pronoyaux migrent vers le centre de l'oeuf. Dans les oeufs mésolécithes ou télolécithes, la fusion des pronoyaux s'effectue au centre de l'aire de cytoplasme actif, soit au pôle animal.
4- activation du métabolisme de l'oeuf et début du développement:
Pour que la fertilisation conduise au développement d'un embryon, des changements doivent se produire dans le cytoplasme de l'oeuf. L'entrée du spermatozoïde met en branle des événements métaboliques préprogrammés, scindés en réponses précoces et tardives.
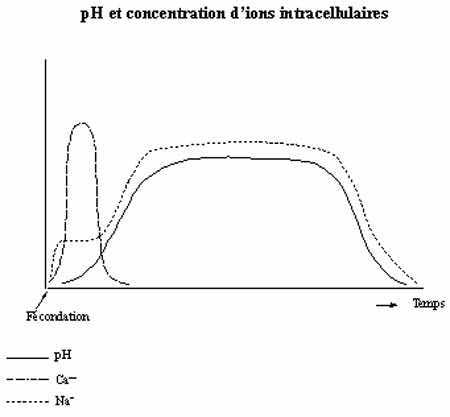
Voir graphique du pH et de la concentration d'ions intracellulaires
Réponses précoces:
-accroissement du pH intracellulaire: résultat de l'influx de Na+ et la perte de H+ amenés par l'activation du blocage de la polyspermie
-modifications de l'activité respiratoire: la libération intracellulaire de Ca++ amenée par le blocage de la polyspermie active une série de réactions métaboliques qui accroissent l'utilisation d'O2 par l'oeuf par l'activation de cytochromes oxydases et l'activation de NAD-kinase, qui catalyse la synthèse de NADP+ à partir de NAD+ et ATP. Le NADP+ est en plus utilisé comme co-enzyme dans la synthèse lipidique, importante pour la construction des membranes en vue de la segmentation prochaine. La fécondation a donc pour effet d'activer l'élimination des déchets métaboliques accumulés dans l'oeuf vierge, qui était plus ou moins en état d'anoxie. Ce flux de Ca++ est essentiel à l'activation du métabolisme de l'oeuf, qui peut d'ailleurs être activé en l'absence de spermatozoïde, par injection de Ca++: c'est la parthénogenèse artificielle.
Réponses tardives:
- Activation de la synthèse d'ADN
- Activation de la synthèse de protéines qui ne dépend pas de la synthèse de nouveaux ARNm mais de ceux mis en réserve dans l'oeuf durant l'ovogenèse. Tous les ARNm préformés ne sont pas traduits simultanément après la fécondation.
- Réorganisation du cytoplasme de l'oeuf. Les morphogènes accumulés durant l'ovogenèse devront être assignés à des cellules-filles spécifiques durant la segmentation. Pour ce faire, ils sont répartis spatialement et de façon spécifique dans l'oeuf peu après la fécondation. De leur organisation spatiale dépendra le développement normal de l'embryon. Le cytoplasme cortical de l'oeuf nouvellement fécondé subit une rotation de 30° par rapport au cytoplasme profond. L'entrée du spermatozoïde dans l'oeuf amène la réorganisation cytoplasmique en dictant le sens de la rotation.
La position du premier plan de segmentation n'est donc pas établie au hasard mais spécifiée par le point d'entrée du spermatozoïde dans l'oeuf. Ce point définit la future face ventrale de l'embryon. La coordination du réarrangement cytoplasmique et du plan de clivage est probablement médiée par les microtubules de l'aster du spermatozoïde. Les centrioles mâles non seulement organisent l'appareil mitotique mais réorganisent le cytoplasme du zygote, maintenant prêt pour le développement d'un animal multicellulaire.
Parthénogenèse:
La parthénogenèse est le mode de reproduction selon lequel une activation spontanée de l'oeuf lui permet de se développer en un nouvel individu sans qu'il y ait fécondation. La parthénogenèse naturelle se rencontre chez plusieurs groupes zoologiques mais survient surtout dans des conditions écologiques particulières. Elle peut se produire chez toutes les classes de Vertébrés, sauf les mammifères. Elle peut être provoquée expérimentalement chez des espèces où la fécondation est la règle, tels l'oursin et des amphibiens. Ces expériences permettent d'analyser les mécanismes de la fécondation normale.
La parthénogenèse naturelle s'accomplit par une des variations sur le thème de la méiose. Par exemple, chez Drosophila mangabeirai un des corps polaires agit comme spermatozoïde et "fertilise" l'ovocyte après la deuxième division méiotique. Chez l'insecte Moraba virgo et le lézard Cnemidophorus uniparens l'oeuf double son nombre de chromosomes avant la méiose; les deux divisions méiotiques restaurent donc la diploïdie. Chez la sauterelle Pycnocelus surinamensis il ne se produit pas de méiose, les oeufs se forment par deux divisions mitotiques. Tous ces cas résultent en femelles diploïdes.
Chez d'autres espèces, la parthénogenèse haploïde est utilisée non comme moyen de reproduction, mais comme mécanisme de détermination sexuelle. Par exemple, chez les hyménoptères (abeilles, guêpes, fourmis) les oeufs non fertilisés, haploïdes, se développent en mâles tandis que les oeufs fertilisés, diploïdes, se développent en femelles. Les mâles haploïdes peuvent produire des spermatozoïdes en abandonnant la première division méiotique, formant ainsi deux spermatozoïdes haploïdes.
La parthénogenèse expérimentale peut être induite par des agents chimiques. Les substances ionophores liant le calcium peuvent entraîner une activation parthénogénétique (chez l'oursin, les amphibiens et le hamster) en provoquant un afflux de Ca++ dans l'ovocyte, afflux qui a le même effet que l'entrée du spermatozoïde à la fécondation normale: achèvement de la méiose, émission des granules corticaux, activation métabolique de l'oeuf. Il est à noter que chez les mammifères, l'embryon parthénote, s'il réussit à s'implanter, voit son développement arrêté à la mi-gestation. La parthénogenèse peut aussi être induite par stimulation traumatique, provoquant le globule polaire à stimuler le développement de l'ovocyte.
Lors de la fécondation normale, le spermatozoïde joue le rôle de stimulus du développement de l'oeuf, par action superficielle analogue à un élément chimique ou à un traumatisme. Bien que non indispensable au rétablissement de la diploïdie et à la réalisation de la segmentation subséquente, il joue néanmoins ces rôles dans la fécondation normale.
L'absence de parthénogenèse naturelle chez les mammifères, l'avortement des parthénotes expérimentaux, de même que différentes expériences de transfert nucléaire tendent à indiquer la nécessité du génome paternel pour le développement normal et total de l'embryon mammalien (Renard et al., Reprod. Nutr. Devel., 28(1988):1541-1554).
Gamétogenèse et fécondation sont deux étapes de la progenèse qui résultent en la formation du zygote, prérequis de l'embryogenèse proprement dite.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}