> Gamétogenèse mâle: spermatogenèse
> Gamétogenèse femelle: ovogenèse
GAMETOGENESE (cours 1)
Gamétogenèse: différenciation des cellules germinales ou reproductrices, les gamètes.L'isogamie est la condition selon laquelle les gamètes des deux sexes sont semblables. Mais chez la plupart des espèces, les deux types de gamètes sont morphologiquement différents; il s'agit d'anisogamie ou hétérogamie:
gamète mâle: spermatozoïde
gamète femelle: oeuf
La gamétogenèse prend le nom de spermatogenèse ou d'ovogenèse, selon le sexe. Elle se produit dans des organes spécialisés, les gonades: gonade mâle: testicule
gonade femelle: ovaire
Les gonades des Vertébrés jouent aussi un rôle endocrinien en renfermant dans leur tissu des cellules endocrines, d'où le terme de glandes génitales.
Chez les espèces dites gonochoriques, spermatogenèse et ovogenèse se déroulent chez des individus différents. C'est le cas de la plupart des Vertébrés, sauf certains poissons et reptiles. Le sexe des individus gonochoriques est déterminé génétiquement dès la fécondation par la nature des chromosomes sexuels, symbolisés par X et Y chez les mammifères et Z et W chez les oiseaux.
- L'un des sexes est homogamétique et ne forme qu'une catégorie de gamètes: la femelle (XX) chez les mammifères et le mâle (ZZ) chez les oiseaux.
- L'autre sexe est hétérogamétique et forme deux catégories de gamètes différents par la nature d'un chromosome sexuel: le mâle (XY) chez les mammifères et la femelle (ZW) chez les oiseaux.
Chez les espèces dites hermaphrodites, un même individu porte les gonades mâle et femelle, comme on le rencontre fréquemment chez les Invertébrés.
- Hermaphrodite spontané: un même individu peut avoir simultanément des activités mâle et femelle, et la fécondation entre partenaires est réciproque. Ex. ver de terre, escargot.
- Hermaphrodite successif: un même individu est successivement à activité mâle puis femelle. Ex. certains mollusques.
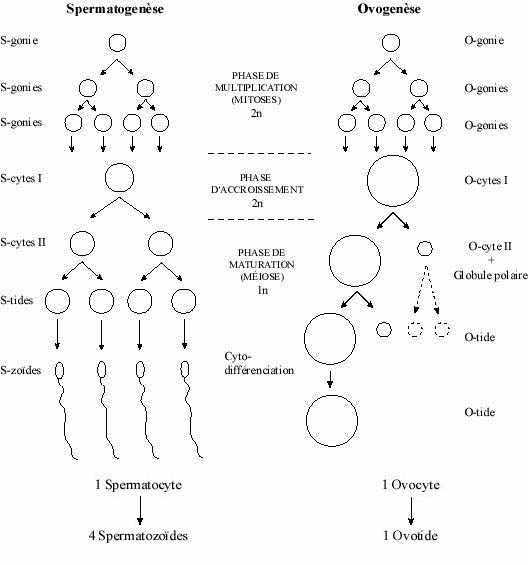
La différenciation des gamètes présente une certaine uniformité dans le règne animal. Ainsi peut on décrire des stades communs à la spermatogenèse et à l'ovogenèse de toute espèce.
1- phase de multiplication: la gamétogenèse commence par une phase de multiplication pendant laquelle les cellules germinales diploïdes, spermatogonies et ovogonies, se divisent par mitoses et augmentent leur nombre.
2- phase d'accroissement: les gonies cessent de se diviser par mitoses et prennent le nom d'auxocytes primaires: spermatocytes I et ovocytes I; leur volume augmente par accroissement du cytoplasme. Les auxocytes I entrent en prophase de la première division méiotique et répliquent leur ADN.3- phase de maturation: marquée par la méiose. Les auxocytes I deviennent haploïdes, auxocytes secondaires: spermatocytes II et ovocytes II, puis, après la 2e division de méiose, spermatides et ovotides. Pendant cette phase se produit aussi une cytodifférenciation conduisant à l'anisogamie et résultant en la formation de gamètes fonctionnels mâle: spermatozoïde, et femelle: oeuf.
GAMETOGENESE MALE: SPERMATOGENESE
Voir schéma sur le système reproducteur mâle (humain)
La spermatogenèse a été décrite pour la première fois dans le testicule du ver nématode Ascaris par Hertwig en 1880.
Chez la plupart des Vertébrés et de nombreux Invertébrés, les tissus séminifères sont organisés en tubules, nombreux et souvent contournés: les tubules séminifères. Ils sont séparés les uns des autres par du tissu conjonctif interstitiel renfermant, entre autre, des cellules à fonction endocrine, sécrétrices d'hormone, les cellules interstitielles de Leydig qui produisent la testostérone. Le testicule est entouré d'une capsule de tissu conjonctif, la tunique albuginée du testicule. Les tubules séminifères convergent vers la sortie du testicule et fusionnent en quelques tubules efférents, puis en un tubule unique, le canal de l'épididyme, long et maintes fois replié sur lui-même. Le canal de l'épididyme forme la structure appelée épididyme qui repose à la surface du testicule. Il se continue en un tube à paroi plus épaisse, le canal déférent, ou canal de Wolff. Les canaux déférents gauche et droit se jettent dans l'urètre, unique. L'urètre traverse le pénis, organe copulateur. Epididyme, canal déférent et urètre constituent les voies génitales mâles que doivent traverser les spermatozoïdes pour aller féconder la gamète femelle.
Un tubule séminifère est fait d'une paroi comprenant un épithélium stratifié souligné d'une membrane basale, elle-même sous-tendue de cellules contractiles appelées cellules péritubulaires ou myoïdes et de tissu conjonctif délicat. L'épithélium est composé de deux types cellulaires:
1- les cellules de la lignée germinale (spermatique), à renouvellement continu et qui se différencient en spermatozoïdes qui seront largués dans la lumière du tubule, et
2- les cellules de Sertoli, de soutien et nourricières des cellules germinales. Elles s'étendent de la base à l'apex de l'épithélium. Elles émettent de nombreux bras cytoplasmiques qui s'insèrent entre les cellules germinales et les entourent mais elles demeurent isolées des cellules germinales par une membrane basale. Leur noyau est volumineux, avec une chromatine diffuse et un gros nucléole, indications d'une activité de synthèse d'ARN, et leur cytoplasme contient des inclusions de réserves: gouttelettes lipidiques, glycogène et phosphatases. Elles phagocytent les cellules germinales qui dégénèrent ainsi que les résidus des spermatozoïdes mûrs (voir plus loin). Dans le testicule foetal, elles sécrètent des hormones anti-mülleriennes (AMH), qui dictent la dégénérescence du canal de Müller (voir Développement du système reproducteur), et la substance SGF: "spermiogenesis growth factor" (voir plus loin).
La spermatogenèse comprend deux étapes, spermatogenèse proprement dite et spermiogenèse.
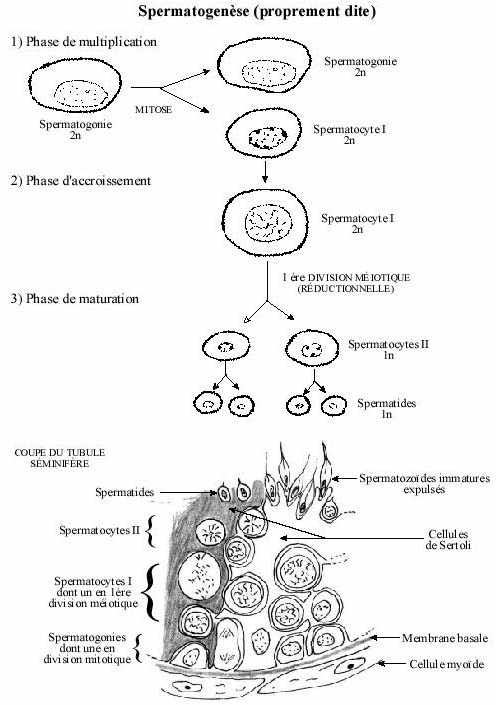
Spermatogenèse (voir schéma)
1- phase de multiplication:
Processus continu commençant dès la vie foetale, il devient très actif à la puberté (début de la maturité sexuelle et de la vie reproductrice) et se poursuit jusqu'à la sénescence. Les spermatogonies, diploïdes, se divisent par mitoses et augmentent leur nombre. Certaines de leurs cellules-filles demeurent cellules-souche à la base de l'épithélium du tubule séminifère; leur chromatine est condensée. D'autres cessent de se diviser et sont repoussées vers l'apex de l'épithélium; leur chromatine est diffuse. C'est à partir de ce moment qu'est calculé le début du cycle spermatogénique (voir plus loin). Ces cellules plus petites sont riches en ribosomes et sont reliées entre elles par des ponts cytoplasmiques (gap junctions). Elles portent maintenant le nom de spermatocytes I. S'entame maintenant la phase suivante.
2- phase d'accroissement:
De brève durée. Les spermatocytes I, diploïdes, répliquent leur ADN (début de la première division méïotique) et accroissent leur volume total. Les spermatocytes issus d'une même spermatogonie restent reliés par des ponts cellulaires permettant l'échange d'informations et assurant la synchronie de leur différenciation.
3- phase de maturation:
Commence à la puberté. La première division méiotique (réductionnelle) des spermatocytes I se termine. Ceux-ci sont maintenant appelés spermatocytes II, haploïdes et de taille deux fois moindre. Cette phase comprend aussi une synthèse active d'ARN dans les autosomes (les chromosomes non sexuels), ARN qui contrôle probablement la différenciation des spermatides. Les spermatocytes II subissent la deuxième division méiotique (méiose équationnelle) et prennent le nom de spermatides, repoussées de plus en plus vers la lumière du tubule séminifère. Ainsi, un spermatocyte I donne naissance à quatre spermatides. Font aussi partie de la phase de maturation les changements morphologiques et biochimiques que subissent les spermatides pour devenir spermatozoïdes. Ces changements constituent la spermiogenèse.
Le cycle spermatogénique est la durée nécessaire à la différenciation d'une spermatogonie devenue post-mitotique (donc prenant le nom de spermatocyte I) en spermatozoïde mûr, incluant la spermiogenèse décrite ci-après. Ce temps est déterminé précisément pour chaque espèce, et à l'intérieur du cycle, chaque étape de différenciation a une durée précise. Par exemple, le cycle spermatogénique de la souris est de 26 jours, celui du rat de 40 jours (dont 12,8 jours dans l'épithélium séminifère) et celui de l'homme de 76 jours.
Dans toute région de l'épithélium séminifère sont superposées des cellules germinales à différentes étapes de la spermatogenèse; une nouvelle génération de spermatogonies commence à se multiplier avant que les cellules de la génération précédente ne soient devenues spermatozoïdes. Des vagues de mitoses se succèdent à un rythme régulier. Toutefois, la vague mitotique n'est pas entamée simultanément sur toute la longueur d'un tubule séminifère donné. Une vague mitotique commence à l'extrémité d'un tubule et se poursuit le long du tubule jusqu'à l'autre extrémité. Ceci explique que des coupes transversales passant par différentes régions d'un même tubule ne présentent pas toutes le même stade de la spermatogenèse (voir Laboratoire 1). Etant donné la durée précise du cycle spermatogénique et celle précise de chaque étape du cycle, en coupe de testicule on peut identifier différents stades de la spermatogenèse, i.e., différentes combinaisons possibles de cellules spermatiques présentes à un niveau donné du tubule vu en coupe transversale. Le nombre de stades identifiables (combinaisons possibles) dépend de la durée du cycle et de la durée de chaque étape, ces paramètres étant spécifiques à une espèce animale. Ainsi, chez le rat, on peut décrire 14 stades du cycle spermatogénique (voir Laboratoire I).
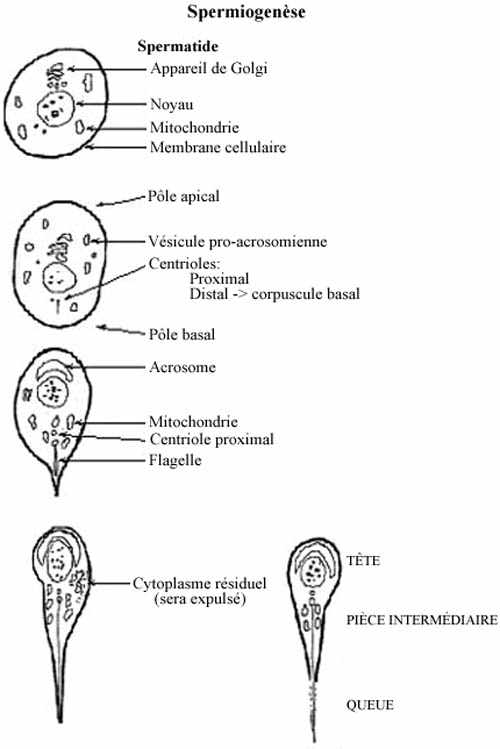
Spermiogenèse: cytodifférenciation de la spermatide en spermatozoïde (voir schéma)
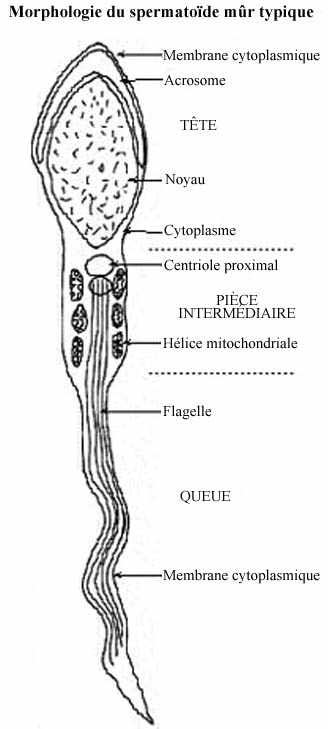
La structure générale des spermatozoïdes est, à quelques exceptions près, uniforme chez toutes les espèces. On retrouve une tête qui comprend le noyau haploïde, coiffé sur sa face apicale de l'acrosome, le tout entouré d'une mince pellicule de cytoplasme; une pièce intermédiaire qui comprend la base du flagelle et l'appareillage énergétique de la cellule; une queue qui comprend surtout un flagelle assurant la motilité du spermatozoïde. Le processus de spermiogenèse débute dans l'épithélium séminifère et se poursuit après que le spermatozoïde en soit expulsé. Une bonne partie des processus de différenciation décrits ici se produit une fois que les spermatozoïdes sont dans l'épididyme.
1- À partir de l'appareil de Golgi se forment des granules glycoprotéiques, les granules pro-acrosomiens, contenus dans des vésicules cytoplasmiques qui migrent vers le pôle apical de la spermatide. Ces vésicules fusionnent et forment l'acrosome, ou capuchon céphalique, qui coiffe la surface apicale du noyau cellulaire. L'acrosome est riche en phospholipides et glycoprotéines, en enzymes lytiques associées à ces molécules (hyaluronidase et hydrolases) et en une enzyme analogue à la trypsine. C'est donc un gros lysosome modifié.
2- Les deux centrioles de la spermatide migrent vers le pôle basal. L'un d'eux, le centriole distal, forme le corpuscule basal, ou cinétosome, à l'origine du flagelle.
3- La taille du noyau se réduit, la chromatine nucléaire se condense et l'acrosome adapte sa forme à celle du noyau, recouvrant environ les deux tiers apicaux de celui-ci.
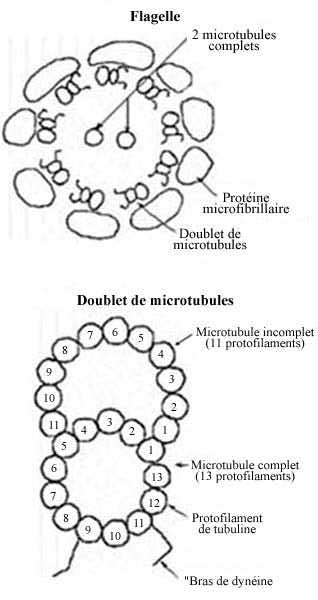
4- Des microtubules ancrés au cinétosome commencent à former le flagelle. Ce dernier se compose de deux microtubules centraux entourés de neuf doublets de microtubules, tous fusionnés au niveau du centriole. Le tout est entouré de microfibrilles.
5- Le cytoplasme, avec le reste des organites, se déplace vers la région basale de la cellule et entoure la partie proximale du flagelle en formation. La forme du noyau et de l'acrosome devient de plus en plus caractéristique de l'espèce. La chromatine nucléaire achève de se condenser. Le flagelle continue de s'allonger. Presque tout le cytoplasme est éliminé avec les organites qu'il renferme (Golgi, ribosomes, etc.) et ce résidu est phagocyté par les cellules de Sertoli.
6- Les mitochondries, regroupées derrière le noyau, se disposent les unes derrière les autres et forment une chaîne enroulée autour de la base du flagelle, dans la pièce intermédiaire; c'est l'hélice mitochondriale.
Les ARN synthétisés durant la phase d'accroissement, avant la méiose (et éliminés avec le cytoplasme résiduel), codaient pour la synthèse de protéines nécessaires à la formation des organites des spermatozoïdes, entre autres les protéines microfibrillaires et microtubulaires, les enzymes de l'acrosome, les protéines nucléaires.
L'ADN haploïde forme 20% du poids sec du spermatozoïde mûr. Les protéines nucléaires autres que les histones sont éliminées; les histones conservées contribuent à la condensation et à la stabilisation de la chromatine, la protégeant contre les altérations qu'elle pourrait subir pendant le passage des spermatozoïdes dans les conduits génitaux mâles, puis femelles (pour les animaux à fécondation interne).
Le spermatozoïde mûr est donc une cellule extrêmement spécialisée, dépourvue de nombreux organites cytoplasmiques, n'ayant conservé que ceux indispensables à sa fonction: le transfert du patrimoine génétique mâle vers l'oeuf de la femelle. Chez les mammifères, il mesure de 40 à 250µm de longueur (53µm chez l'homme) et son volume est considérablement réduit, comparé à celui de l'oeuf. Par exemple, le spermatozoïde du taureau, avec un volume de 30µm3, n'atteint que le 1/20 000 du volume de l'oeuf de la vache (oeuf pourtant petit, relativement - voir chapitre suivant). Voir la morphologie de différents spermatozoïdes
La motilité du spermatozoïde est assurée par la pièce intermédiaire et le flagelle. La majeure partie du flagelle, l'axonème, consiste en deux microtubules centraux entourés de neuf doublets de microtubules. Dans chaque doublet, un seul microtubule est complet, avec 13 protofilaments, l'autre ayant la forme d'un C, avec 11 protofilaments. Les protofilaments sont composés exclusivement de la protéine dimérique tubuline. La dynéine, une protéine attachée aux microtubules, hydrolyse les molécules d'ATP et convertit l'énergie chimique libérée en énergie mécanique qui propulse le spermatozoïde. L'hélice mitochondriale de la pièce intermédiaire possède l'équipement nécessaire aux phosphorilations oxydatives apportant l'énergie nécessaire aux battements du flagelle et à la survie des spermatozoïdes. Les sources d'énergie sont fournies soit par des éléments externes, tel le fructose du liquide séminal, soit par des réserves endocellulaires, comme des phospholipides. Elles sont dégradées par les processus de la glycolyse en l'absence d'oxygène, avec formation d'acide pyruvique et lactique. En présence d'O2, les enzymes mitochondriales achèvent leur dégradation complète avec formation de CO2 et H2O. Ce mécanisme permet aux spermatozoïdes d'effectuer de longs parcours. Chez l'homme, ils peuvent se déplacer à une vitesse de 2-3µm/min. à 37°. La motilité est acquise dans l'épididyme, sous l'effet des sécrétions fournies par cette glande, additionnées des sécrétions des vésicules séminales et de la prostate rejetées dans le canal déférent. (Chez certaines espèces, Ascaris par exemple, les spermatozoïdes se meuvent par mouvements amiboïdes, grâce à des pseudopodes.) Au sortir du testicule, les spermatozoïdes n'ont pas encore la capacité de féconder l'oeuf. Pour ce faire, il leur faut subir encore deux séries de transformation: l'acquisition du pouvoir fécondant dans les voies génitales mâles, et la capacitation dans les voies femelles (chez les animaux à fécondation interne).
Le pouvoir fécondant est acquis lors de la traversée de l'épididyme, grâce à ses sécrétions. L'hélice mitochondriale achève alors sa mise en place et l'acrosome adopte sa forme définitive. Des sécrétions glycoprotéiques de l'épididyme se déposent à la surface membranaire du spermatozoïde et contribuent à la stabilisation de la membrane: elles masquent les sites antigéniques à la surface du spermatozoïde, lui assurant une impunité contre d'éventuelles agressions dans les voies femelles, et inhibent les enzymes de l'acrosome, évitant que celles-ci ne s'attaquent aux cellules des voies mâles ou femelles. Chez les animaux aquatiques à fécondation externe, motilité et pouvoir fécondant sont acquis au contact de l'eau.
La capacitation se produit pendant le séjour de plusieurs heures qu'effectuent les spermatozoïdes dans les voies génitales femelles (chez les animaux à fécondation interne), et consiste en modifications des propriétés de leur membrane cytoplasmique. Par l'action d'enzymes protéolytiques du liquide utérin, la membrane est débarassée des sécrétions de l'épididyme qui lui étaient accolées; ses sites antigéniques sont démasqués et l'inhibition des enzymes acrosomales est levée. Il se produit une augmentation du métabolisme cellulaire. La capacitation est facilitée en période d'ovulation car le liquide utérin est enrichi en enzymes protéolytiques. La capacitation est réversible si les spermatozoïdes sont replacés dans le liquide séminal.
La durée de vie des spermatozoïdes dans les voies femelles varie selon les espèces. Elle ne dépasse guère quelques jours chez les mammifères (sauf les chauves-souris dont les spermatozoïdes ont une plus longue viabilité). Notons un cas extrême, celui de l'abeille reine, chez qui ils peuvent survivre deux ou trois ans dans la spermathèque.
L'activité mitotique de l'épithélium séminifère est très intense et dure pendant toute la vie reproductrice. Le lapin, par exemple, produit 80 000 spermatozoïdes/ minute. Chez l'homme, le volume de l'éjaculat est de 3 à 3,5cc et renferme de 60 000 000 à 120 000 000 spermatozoïdes/cc; environ 300 000 000 spermatozoïdes sont donc émis à chaque éjaculation.
Le taureau éjacule 4cc de sperme qui renferme 1 000 000 000 de spermatozoïdes/cc
Le chien éjacule 6cc de sperme qui renferme 200 000 000 de spermatozoïdes/cc
L'étalon éjacule 70cc de sperme qui renferme 100 000 000 de spermatozoïdes/cc
Le verrat éjacule 250cc de sperme qui renferme 200 000 000 de spermatozoïdes/cc
Le liquide séminal est le véhicule liquide des spermatozoïdes. Sa production commence dans les tubules séminifères (fluide tubulaire), par le transport de plasma à partir des capillaires du tissu conjonctif interstitiel du testicule jusque dans la lumière des tubules. Le plasma exsudé doit donc traverser l'épithélium séminifère et, ce faisant, sa composition est modifiée par des sécrétions des cellules de Sertoli. Le fluide et les spermatozoïdes sont amenés dans l'épididyme. Les cellules sécrétices de la paroi épithéliale des tubules de l'épididyme ajoutent considérablement au fluide séminal, de même que les cellules des autres glandes des voies génitales mâles, jusqu'à l'obtention du fluide final qui sera éjaculé: le sperme.
Contrôle endocrinien de la spermatogenèse: deux hormones glycoprotéiques de structure voisine, élaborées par l'hypophyse antérieure:
FSH: follicle-stimulating hormone, joue un rôle dans la croissance testiculaire et le déclenchement de la spermatogenèse à la puberté, en plus de stimuler l'activité mitotique de l'épithélium séminifère.
ICSH: interstitial cell stimulating hormone, stimule la synthèse d'hormones androgènes, notamment la testostérone élaborée par les cellules interstitielles, et agit sur l'apparition des caractères sexuels secondaires. Elle stimule aussi l'activité mitotique des spermatogonies et agit sur les cellules péritubulaires en contrôlant la libération des spermatozoïdes dans la lumière des tubules séminifères. C'est l'analogue de la LH femelle.
Les hormones hypophysaires sont elles-mêmes sous le contrôle d'hormones déclenchantes (releasing hormones), FSH-RH et ICSH-RH, sécrétées par l'hypothalamus. Par voie hypothalamique (système limbique), les stimuli externes peuvent agir sur l'activité spermatogénique. Par exemple, la lumière chez les oiseaux et les mammifères peut provoquer une décharge d'ICSH qui résulte en un pic de testostérone. Les androgènes, en concentration suffisante dans le sang, ont un effet rétro-inhibiteur sur l'activité de l'hypothalamus.
Les androgènes, notamment la testostérone, élaborés pas les cellules interstitielles de Leydig contrôlent en partie le développement des caractères sexuels gonochoriques et somatiques chez l'embryon. Chez l'adulte, tel que mentionné, la testostérone augmente l'activité de l'épithélium germinal, donc la spermatogenèse, par action directe (par diffusion locale plutôt que par la circulation sanguine) sur les tubules séminifères. De plus, elle agit sur les cellules de Sertoli, pour que celles-ci créent l'environnement optimal à la fonction spermatogénique, et sur la vascularisation. Les hormones stéroïdes sécrétées par les gonades dérivent du cholestérol et en conservent la structure polycyclique.
Un facteur de croissance sécrété par les cellules de Sertoli, le SGF (spermiogenesis growth factor) stimule la prolifération cellulaire dans l'épithélium séminifère et la synthèse de protéines nécessaires à la spermatogenèse. Durant l'ontogenèse (étudiée chez la souris), le SGF déclenche l'activité mitotique des cellules de Sertoli, des cellules péritubulaires (myoïdes) et des cellules interstitielles (de Leydig), en plus de stimuler la formation de vaisseaux sanguins dans le testicule. Il existe donc une interaction complexe entre les cellules interstitielles, les cellules de Sertoli, les cellules péritubulaires, les cellules germinales et la vascularisation. Voir tableau de construction des principales structures du spermatozoïdes.
D'autres facteurs affectent la spermatogenèse:
température: une fois produits, les spermatozoïdes sont amenés dans l'épididyme où ils sont emmagasinés pendant un certain temps. Dans l'épididyme, selon que l'espèce possède des testicules externes (dans le scrotum) ou en position abdominale, les spermatozoïdes se retrouvent à température inférieure de quelques degrés à la température corporelle. À température corporelle, le sperme serait moins viable. Cette théorie est de plus en plus contestée, à cause notamment du plus grand nombre d'espèces ayant des testicules internes (voir p. 120).
nutrition: une carence en vitamines A et E et en acides gras résulte en une diminution de la spermatogenèse.
radiations ionisantes: les spermatogonies, qui se divisent très activement, y sont très sensibles. Une exposition à des radiations au dessus d'un certain seuil peut résulter en stérilité définitive.
Les pesticides ont aussi une action stérilisante.
Les anomalies de la spermatogenèse, résultant en une structure anormale du spermatozoïde qui perd son pouvoir fécondant, sont plus fréquentes chez les espèces à faible taux de fécondité. Le spermatozoïde peut être bicéphale, microcéphale, biflagellé, etc. Généralement, les spermatozoïdes porteurs de chromosomes anormaux dégénèrent. Chez le mulet, résultat du croisement d'un âne et d'une jument, l'appariement des chromosomes se fait difficilement et les gamètes dégénèrent au stade spermatocyte I. Pour que le sperme soit normalement fécond, il faut que moins de 20% des spermatozoïdes soient anormaux. Lorsque plus de 50% le sont, on parle de tératospermie. La stérilité peut aussi être due à une émission insuffisante de sperme, à un nombre insuffisant de spermatozoïdes dans le sperme (oligospermie), à l'absence de spermatozoïdes dans le sperme (azoospermie) ou à des spermatozoïdes insuffisamment mobiles (athénospermie).
GAMETOGENESE FEMELLE: OVOGENESE (cours 2)
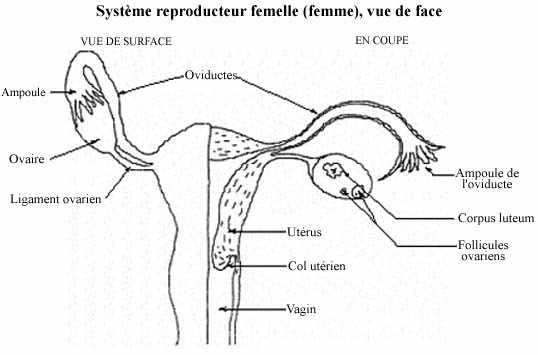
Voir schéma sur le système reproducteur femelle (humain)
La formation des cellules reproductrices femelles, les oeufs, se déroule dans les gonades femelles, les ovaires. Les cellules de la lignée germinale ne sont pas organisées dans des tubules comme chez le mâle, mais dispersées dans le stroma conjonctif de l'ovaire. Chaque cellule germinale est entourée de cellules de soutien appelées cellules folliculaires; la cellule germinale et les cellules folliculaires qui y sont associées forment ensemble le follicule ovarien.
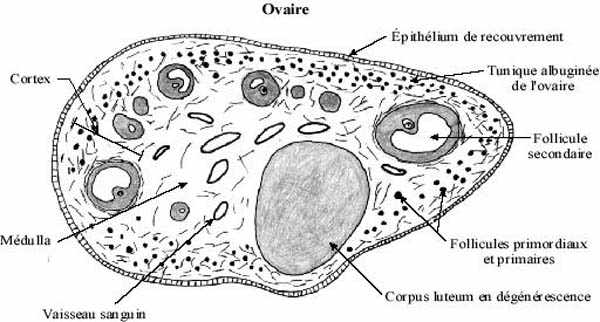
L'ovaire comprend deux régions histologiques, la médulla centrale et le cortex. Le cortex renferme les follicules séparés les uns des autres par du tissu conjonctif délicat et des petits vaisseaux sanguins, la médulla renferme du tissu conjonctif et des vaisseaux plus gros. Les premières étapes de l'ovogenèse se produisent dans l'ovaire embryonnaire.
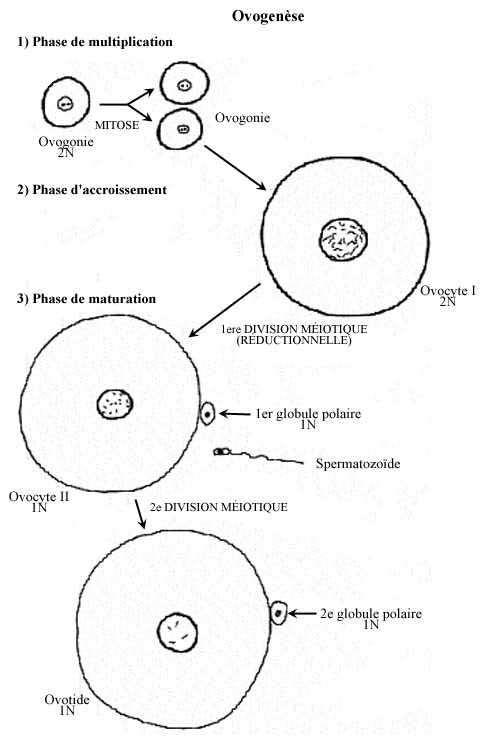
1- phase de multiplication:
Dans l'ovaire embryonnaire des Vertébrés, les cellules germinales se multiplient activement et portent le nom d'ovogonies. Cellules diploïdes arrondies, d'aspect indifférencié et de grande taille, leur noyau renferme un ou deux nucléoles et une chromatine finement granulaire, et leur cytoplasme possède peu d'organites: quelques mitochondries, peu de réticulum endoplasmique et un appareil de Golgi réduit. Les ovogonies issues d'une même cellule-souche sont reliées par des ponts cytoplasmiques et leurs divisions sont synchrones.
Leur activité mitotique est limitée dans le temps chez de nombreuses espèces. Chez les amphibiens et les téléostéens, l'activité mitotique des ovogonies est saisonnière: les mitoses surviennent après chaque ponte et reconstituent un lot d'ovogonies qui, après croissance, fourniront une ponte ultérieure; des nids d'ovogonies dormantes subsistent en permanence dans les ovaires. Chez les autres Vertébrés, les mitoses cessent avant la fin de la vie embryonnaire ou après la naissance. Par exemple, chez le poussin les divisions mitotiques cessent 4 à 8 jours après l'éclosion. Chez l'humain, les ovogonies cessent de se diviser à la 15e semaine de vie utérine; elles sont alors au nombre d'environ 4 millions.
2- phase d'accroissement:
Plusieurs ovogonies dégénèrent; environ la moitié seulement des 4 millions (chez l'humain) entame la phase d'accroissement: on les appelle ovocytes I. Les ovocytes I ne sont plus reliés par des ponts cytoplasmiques. La longue phase d'accroissement peut se scinder en deux sous-phases. Une sous-phase de petit accroissement s'étend de la vie foetale à la puberté (âge de la maturité sexuelle) ou même plus tard (voir plus loin) et voit la dégénérescence de plusieurs ovocytes I. Par exemple, chez l'humain il en reste environ 500 000 à la naissance et 7 000 à la puberté; de ceux-ci environ 500 seulement termineront la seconde sous-phase d'accroissement, celle de grand accroissement.
L'ovocyte I des mammifères accoît son diamètre de 30µm à 140µm, soit d'un facteur d'environ 4.5; celui de la grenouille d'un facteur de 27 000. La phase d'accroissement se caractérise par des activités de synthèses intenses: ADN, ARN et protéines, et par l'accumulation de différents matériaux exogènes, en vue du développement de l'embryon. L'accumulation de vitellus se produit pendant la sous-phase de grand accroissement et son importance varie selon les espèces.
Au début de la phase d'accroissement, les ovocytes I, diploïdes, entrent en prophase de la première division méiotique. Le noyau est volumineux, avec de gros nucléoles. La première division de méiose se poursuit par une spiralisation des chromosomes au stade leptotène et leur appariement par éléments homologues au stade zygotène. Ils ont alors un aspect bien particulier selon les espèces: en écouvillon chez les amphibiens, en filament épais chez l'humain (noyau dictyé). Au stade pachytène apparaissent les complexes synaptosémaux qui représentent les aires d'appariement des chromosones homologues. Dans le cytoplasme des ovocytes I, les organites se rassemblent en grande partie dans une aire limitée, voisine du noyau, anciennement appelée noyau vitellin de Balbiani. Le stade diplotène est atteint peu après la naissance chez la plupart des Vertébrés, les complexes synaptosémaux disparaissent, les chromosomes homologues n'étant plus appariés qu'aux chiasma (crossing over). L'ovocyte I reste au stade diplotène jusqu'à la phase de maturation, qui débute à la puberté ou plus tard.
3- phase de maturation:
Caractérisée par la poursuite de la méiose, elle commence à la puberté ou plus tard dans la vie adulte, jusqu'à la ménopause. Donc, tous les ovocytes I n'entament pas cette phase en même temps; la maturation se produit selon un cycle précis et n'implique que quelques ovocytes à la fois. Ainsi, dans l'ovaire d'une femelle amniote sexuellement mûre, on trouve des ovocytes I à l'état "dormant" et d'autres en processus actif de maturation. La première division méiotique résulte en deux cellules haploïdes de taille très inégale: un gros ovocyte II et un tout petit globule polaire, destiné à dégénérer.
Chez l'humain, la ponte ovulaire, ou ovulation, se produit juste après la première division méiotique. C'est donc un ovocyte II qui est largué par l'ovaire et capté par le pavillon de l'oviducte. Chez d'autres espèces, l'ovulation peut se produire un peu plus tôt ou un peu plus tard dans le cycle méiotique. Chez l'humain, généralement un seul ovocyte II est émis tous les 28 jours environ, depuis la puberté jusqu'à la ménopause, par chacun des ovaires alternativement. Le nombre total de gamètes produits par une femme pendant sa vie entière est de l'ordre de 500 au maximum. Que différent de l'homme!
L'ovocyte II entame la seconde division méiotique, qui reste bloquée en métaphase (blocage dû à l'action de la kinase sérine-thréonine de la protéine Mos ainsi qu'à l'activation du complexe cyclin-p34cdc2). Elle ne peut s'achever qu'en présence du spermatozoïde, qui produit un effet activateur en dégradant le complexe. L'ovocyte II qui achève la seconde division méiotique donne naissance à deux cellules haploïdes, elles aussi de taille fort différente: un gros ovotide et un petit (deuxième) globule polaire qui dégénérera. Chez certaines espèces, le spermatozoïde pénètre l'oeuf avant même la fin de la première division méiotique, alors que l'oeuf est encore diploïde.
Synthèses pendant l'ovogenèse
Alors que le spermatozoïde n'a conservé que le strict nécessaire à la fécondation, l'oeuf, au contraire, conserve tout son cytoplasme et en emmagasine même davantage par des synthèses et par la capture de substances exogènes. C'est parce qu'il doit contenir le matériel nécessaire au début du développement de l'embryon. La rapidité et l'autonomie des premiers stades de développement dépendent de la quantité de substances accumulées pendant l'ovogenèse. Ceci s'éclaircira en étudiant le développement embryonnaire. Quelles sont ces synthèses effectuées durant la formation de l'oeuf?
1) réplication de l'ADN:
L'ovocyte I réplique d'abord son ADN nucléaire (chromosomique) puis son ADN cytoplasmique (mitochondrial). À la fin de l'ovogenèse, il y a 100 fois plus d'ADN mitochondrial que d'ADN nucléaire.
2) synthèses d'ARN:
ARNt: Les ARNt sont synthétisés tout au long de l'ovogenèse et s'accumulent, pour la plupart, dans le cytoplasme sous forme de particules ribonucléiques qui contiennent aussi des 5S ribosomaux: les informosomes. Un oeuf mûr d'amphibien, par exemple, contient 60,000 X plus d'ARNt qu'une cellule somatique. Ils sont associés aux acides aminés correspondants et à des aminoacyl-t-ARN synthétases. Ces ribonucléoprotéines sont stables et les ARNt seront utilisés pendant la segmentation de l'embryon.
ARNm et ARNn: Les ARN nucléaires sont aussi appelés ARN hétérogènes à cause de leur coefficient de sédimentation variable. Avec les ARNm ils constituent les ARN informationnels. Ils sont synthétisés au stade diplotène sur les boucles des chromosomes écouvillons (voir ci-bas), durant la sous-phase de petit accroissement.
ARNr: L'assemblage des ARN dans les ribosomes se fait pendant la sous-phase de grand accroissement de l'ovocyte; la réserve de ribosomes accumulée pendant l'ovogenèse est stable et servira au début de la vie embryonnaire. Elle suffira à assurer les synthèses protéiques en début de développement, jusqu'à ce que les synthèses d'ARN à partir du génome de l'embryon soient opérantes et suffisantes. Chez les amphibiens, par exemple, les ARNr maternels sont fonctionnels jusqu'à des stades larvaires avancés, ce qui n'est pas le cas chez les mammifères.
Les chromosomes écouvillons se retrouvent dans les ovocytes au stade diplotène chez tous les groupes zoologiques mais se voient particulièrement bien chez les amphibiens à cause de la grande taille de leurs chromosomes. Les redondances géniques sont nombreuses.
Chaque chromosome d'une paire d'homoloques comporte deux chromatides dont la chromatine est condensée à certains niveaux: les chromomères. L'activité transcriptionnelle y est nulle au stade diplotène. À d'autres niveaux la chromatine est décondensée en boucles symétriques d'environ 50nm; ce sont des sites de transcription génique pendant l'ovogenèse. Ces sites actifs représentent 5 à 10% de la longueur totale des chromosomes. Le chromosome écouvillon n'est pas une structure statique mais dynamique: les boucles se déplacent le long des chromatides. Donc, c'est plus que 5 ou 10% des gènes qui sont transcrits pendant l'ovogenèse. La position et la taille des boucles d'un chromosome écouvillon sont caractéristiques de l'espèce. Une grande partie des ARN synthétisés par les chromosomes écouvillons consiste en ARN informationnels, synthèse qui commence pendant la sous-phase de petit accroissement. 90% de l'ARN informationnel est répétitif. Les espèces moléculaires sont donc présentes en plusieurs milliers d'exemplaires. Un grand nombre de gènes sont transcrits dans l'ovocyte.
Une portion des ARN se dégrade durant l'ovogenèse, notamment des ARN hétérogènes, mais la plupart sont stables. Une façon de stabiliser les ARNm est la poly-adénylation. Les acides poly-adényliques sont eux-mêmes très stables et ils stabilisent les ARNm auxquels ils sont liés: ARNm-poly-A. Si les ARN étaient libérés des complexes qui les séquestrent, ils pourraient être traduits; il est donc important qu'ils soient séquestrés pendant l'ovogenèse pour n'être utilisés que pendant les premiers stades embryonnaires.
Dans les petits oeufs alécithes (voir plus loin), les ARN sont synthétisés en moindre quantité et consistent surtout en ARNt et ARNr.
3) Synthèses de protéines:
Synthétisées pendant la phase de grand accroissement, elles consistent principalement en:
histones: protéines qui entrent en quantité égale à l'ADN dans la composition de la chromatine, les histones synthétisées durant l'ovogenèse serviront notablement durant la segmentation de l'embryon, quand le nombre de cellules augmente exponentiellement. Bien que les gènes codant pour les histones soient répétitifs, leur transcription et leur traduction par l'embryon durant sa segmentation ne seraient suffisamment rapides; il faut donc les accumuler durant l'ovogenèse.
tubulines: protéines qui entrent notamment dans la composition des microtubules, les tubulines synthétisées et emmagasinées pendant l'ovogenèse serviront surtout à former les fuseaux mitotiques pour la segmentation de l'embryon. La transcription et la traduction des tubulines ne pourraient s'effectuer suffisamment rapidement par les cellules embryonnaires qui se divisent rapidement; elles doivent donc elles aussi être accumulées pendant l'ovogenèse.
actine: pour la cytocinèse des cellules embryonnaires
protéines ribosomales: qui entrent dans la composition des ribosomes, sont abondantes vue la quantité importante de ribosomes formés durant l'ovogenèse. Les ribosomes servent pendant la synthèse des protéines.
ADN- et ARN-polymérases: abondantes.
Certaines des protéines synthétisées pendant l'ovogenèse sont fondamentales pour le développement de l'embryon. Une mutation les affectant peut arrêter le développement.
4) Synthèses de facteurs morphogènes:
Ils dirigeront la différenciation cellulaire de l'embryon; ils sont distribués différentiellement aux cellules du jeune embryon.
Vitellus:
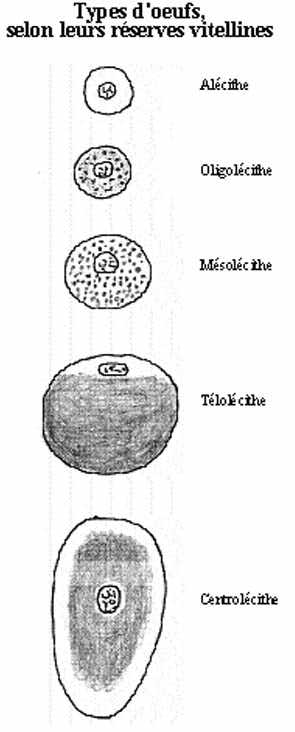
Le vitellus n'est pas une substance chimique bien définie. Il comprend des protéines, phospholipides, graisses neutres et glycogène. Le vitellus est assemblé dans l'ovocyte mais ses précurseurs sont synthétisés ailleurs. Protéines et phospholipides proviennent du foie et passent dans la circulation sanguine, atteignent la thèque interne où ils sont rejetés hors des capillaires sanguins, dans l'espace intercellulaire d'où ils sont captés par les cellules folliculaires, puis transférés à l'ovocyte. Le vitellus servira de réserve nutritive à l'embryon jusqu'à ce qu'il puisse se nourrir par lui-même. Son importance varie selon les groupes zoologiques. La localisation du vitellus dans le cytoplasme varie parmi les groupes dont les oeufs en contiennent beaucoup. Quantité et localisation du vitellus affecteront la fécondation et le développement embryonnaire. Ainsi, selon la quantité et la localisation cytoplasmique du vitellus, il est possible de décrire cinq grands types d'oeufs:
alécithes: n'accumulent pas de vitellus, tels ceux des mammifères euthériens.
oligolécithes: accumulent peu de réserves vitellines, tels ceux des oursins.
isolécithes: les oeufs alécithes et oligolécithes sont aussi dits isolécithes: qui accumulent peu ou pas de vitellus
mésolécithes: accumulent une certaine quantité de vitellus, initialement distribué uniformément dans le cytoplasme (homolécithes) mais devenant ségrégé du cytoplasme actif à la fécondation (hétérolécithe), tels ceux des amphibiens et de certains poissons.
télolécithes: accumulent une très grande quantité de vitellus qui est ségrégé, en masse compacte, du reste du cytoplasme. Ce dernier prend le nom de cytoplasme actif, réduit à un petit disque à la surface du vitellus. Ces oeufs se retrouvent chez les mollusques bivalves, les céphalopodes, les poissons téléostéens, les reptiles et les oiseaux.
centrolécithes: les oeufs des insectes accumulent un abondant vitellus qui forme une masse compacte au centre de la cellule. Le noyau est au centre du vitellus et le cytoplasme actif est périphérique.
Les oeufs mûrs sont généralement sphériques; ils sont allongés, ovoïdes, chez certains animaux: les insectes, certains poissons.
Développement du follicule ovarien mammalien
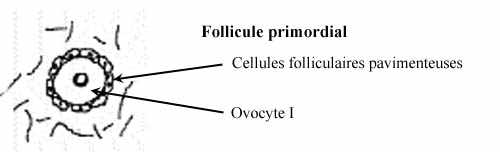
Follicule primordial
Chez l'embryon, les ovogonies sont des cellules arrondies et indifférenciées, de diamètre d'environ 20µm, retrouvées en groupes dans le cortex ovarien, particulièrement à la périphérie du cortex. Par la suite, l'ovocyte I, d'environ 30µm de diamètre après la sous-phase de petit accroissement et bloqué au stade diplotène de la première division méiotique, s'entoure d'une assise de petites cellules somatiques pavimenteuses, reliées par des ponts cytoplasmiques et desmosomes: les cellules folliculaires. Ovocyte I et épithélium folliculaire pavimenteux forment ensemble le follicule primordial. Plusieurs d'entre eux dégénèrent.
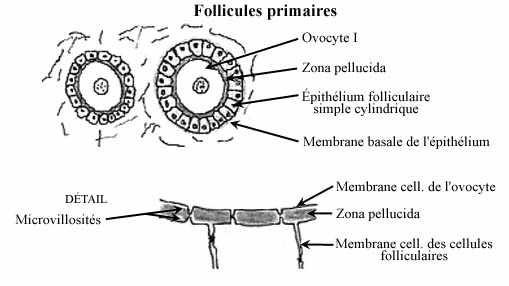
Follicule primaire (schéma)
À partir de la puberté et au premier jour de chaque cycle sexuel, un petit nombre d'ovocytes primaires s'engage dans la sous-phase de grand accroissement puis dans la phase de maturation, tel que décrit ci-haut. Les cellules folliculaires autour d'un ovocyte I deviennent cuboïdales, puis cylindriques. L'ovocyte I sécrète une membrane basale qui devient épaisse et facilement identifiable, la zona pellucida; cette sécrétion glycoprotéique entoure l'ovocyte, le séparant de l'épithélium folliculaire, et agit comme barrière contre la pénétration de substances de haut poids moléculaire, comme des protéines et des polysaccharides. Les membranes cytoplasmiques de l'ovocyte et des cellules folliculaires ne sont pas lisses; elles décrivent de nombreuses petites digitations, genre de microvillosités, qui pénètrent dans la zone pellucide et se touchent l'une l'autre; elles donnent une apparence striée à la zone pellucide (autrefois appellée zona radiata). Les microvillosités permettent d'augmenter considérablement la surface membranaire (celle de l'ovocyte de grenouille est augmentée d'un facteur de 35) et sont l'indice de transports membranaires intensifs. Par pinocytose, elles permettent les échanges de substances de haut poids moléculaire que la zone pellucide empêche de traverser. Les substances exogènes captées par l'ovocyte passent donc d'abord par les cellules folliculaires, qui agissent comme filtre. Le transfert de substances s'effectue moindrement en sens inverse (de l'ovocyte vers les cellules folliculaires). Les cellules folliculaires sécrètent une membrane basale à leur face basale, moins épaisse que la zone pellucide. Les cellules et fibres du tissu conjonctif entourant le follicule se disposent de façon concentrique autour de lui: ébauche de la thèque.
Le terme de zone pellucide s'emploie surtout chez les mammifères; on parle d'enveloppe vitelline chez les autres animaux.
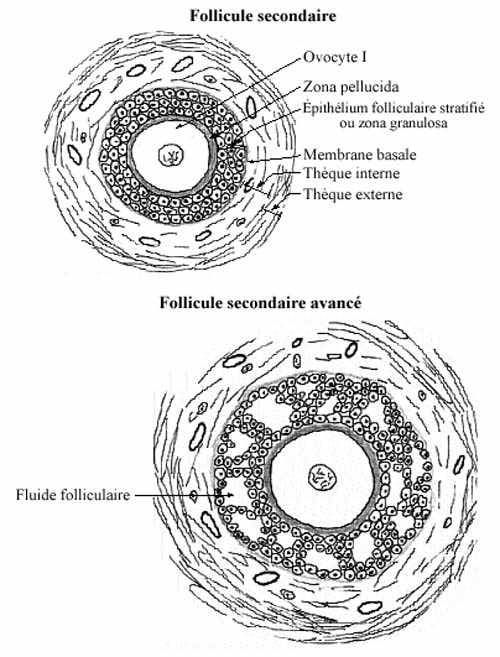
Follicule secondaire (schéma)
L'ovocyte I continue d'accroître son volume. Les cellules folliculaires se divisent par mitoses, formant un épithélium bi- puis pluri-stratifié autour de l'ovocyte, épithélium entouré d'une membrane basale. Elles sont reliées par des ponts cytoplasmiques permettant l'échange de substances de faible poids moléculaire. À cause de la petite taille des cellules, donc du taux nucléoplasmique élevé, l'épithélium folliculaire a une apparence granulaire et porte le nom de zona granulosa. Comme tout épithélium, il est invasculaire; ses cellules, comme l'ovocyte, sont nourries par diffusion à partir des capillaires de la thèque.
La thèque s'épaissit et se subdivise en deux zones: thèque interne, davantage cellulaire que fibreuse, très vascularisée et dont les cellules se différencient en cellules endocrines sécrétrices d'oestrogènes, et thèque externe, plus fibreuse que cellulaire, moins vascularisée, elle a l'aspect habituel du tissu conjonctif, celui-ci arrangé de façon concentrique autour du follicule. Le fluide (liqueur) folliculaire est un sérum qui diffuse à partir des capillaires avoisinants et s'accumule entre les cellules folliculaires, créant ainsi des petits lacs entre elles. Il est riche en oestrogènes, en partie sécrétés par les cellules de la thèque interne. Ovocyte et sa membrane basale, zona granulosa, sa membrane basale et le fluide folliculaire, ainsi que les thèques interne et externe forment ensemble le follicule secondaire. Plusieurs follicules secondaires dégénèrent.
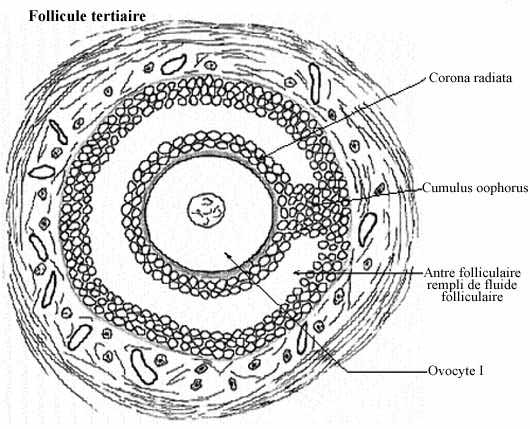
Follicule tertiaire (schéma)
De plus en plus de fluide folliculaire diffuse à partir des capillaires de la thèque interne vers la zone granulaire; les lacs deviennent de plus en plus grands et fusionnent en un bassin unique, l'antre folliculaire. L'antre devient tellement volumineux qu'il forme la plus grande partie du follicule. Il se trouve à diviser la zone granulaire en deux régions: une région interne qui entoure l'ovocyte et sa zone pellucide, de quelques assises cellulaires d'épaisseur et appelée corona radiata, et, autour de l'antre, le reste de la zone granulaire. Cette dernière région demeure rattachée à la couronne radiaire par un pont de cellules folliculaires appelé cumulus oophorus. Pendant ce temps, l'ovocyte I accroît toujours son volume, étant en processus actif de synthèses. Le follicule ovarien à ce stade de développement est appelé follicule tertiaire par certains auteurs et follicule secondaire avancé par d'autres. Plusieurs d'entre eux dégénèrent, phénomène appelée atrésie. Les follicules avancés qui dégénèrent portent le nom de follicules atrésiques.
Follicule mûr ou follicule de De Graaf
Le follicule continue son développement. L'ovocyte I cesse de synthétiser ARN et protéines, les chromosomes se condensent et la diacinèse commence; la première division méiotique s'achève et le premier globule polaire est émis; il demeure accollé contre le gros ovocyte II sous la zone pellucide. Le follicule mûr est tellement gros qu'il fait saillie à la surface de l'ovaire. Chez l'humain, il atteint de 10 à 15mm de diamètre.
Ovulation ou ponte ovulaire
L'accroissement volumétrique du fluide folliculaire dans l'antre exerce une pression considérable sur le tissu folliculaire et entraîne sa rupture à la surface de l'ovaire, rupture qui est également sous le contrôle de l'hormone hypophysaire lutéinisante. Le cumulus oophorus et la zone granulaire se brisent, les thèques cèdent et l'ovocyte, sa zone pellucide et sa couronne radiaire, ensemble l'ovule, sont relâchés dans la cavité pelvienne. Le fluide folliculaire est aussi expulsé. L'ovaire étant recouvert par l'ampoule de l'oviducte (les deux n'étant pas continus mais apposés), l'ovule émis est donc facilement capté par l'ampoule et s'engage dans l'oviducte qui le transporte vers l'utérus.
Ce qui reste du follicule mûr dans l'ovaire, le follicule déhiscent, se transforme en corpus luteum (corps jaune): les capillaires sanguins de la thèque interne envahissent l'espace entre les cellules folliculaires restantes qui, sous l'effet de FSH et LH circulantes, se transforment en cellules endocrines sécrétant progestérones et oestrogènes. Ces hormones gagnent la circulation et préparent la paroi utérine à une nidation éventuelle. S'il y a fécondation le corps jaune est maintenu par l'action d'hormones, sinon il dégénère et une cicatrice se forme, le corpus albicans (corps blanc).
contrôle hormonal de l'ovogenèse
À partir de la puberté, l'hypothalamus sécrète des hormones déclenchantes (releasing hormones - RH) qui stimulent l'activité de l'hypophyse antérieure. Celle-ci, en réponse, élabore deux hormones glycoprotéiques gonadotrophes de structure assez voisine:
FSH: follicle stimulating hormone, active la croissance des follicules ovariens, dont la sécrétion d'oestrogènes par les cellules de la thèque interne. L'atrésie folliculaire est due au fait que le taux de FSH circulante ne suffit pas à assurer la croissance de tous les follicules engagés dans la phase de maturation durant un cycle donné. Seulement quelques follicules, généralement un seul chez l'humain, complètent la maturation. La FSH stimule ensuite la formation du corps jaune.
LH: luteinizing hormone, provoque la rupture du follicule mûr et l'ovulation en plus de déclencher la poursuite de la première division méiotique de l'ovocyte I. La LH stimule la transformation du follicule déhiscent en corps jaune.
Un équilibre doit exister entre les deux hormones: le contrôle de la phase finale de croissance du follicule nécessite un maximum de FSH en présence de LH. Ces hormones déterminent aussi le nombre de follicules qui mûrissent simultanément et qui dégénèrent.
Le cycle sexuel de la femelle peut être divisé en plusieurs phases:
phase folliculaire:
Au début du cycle le taux des hormones stéroïdes sexuelles circulantes est faible, ayant pour effet de stimuler la production de FSH-RH par l'hypothalamus, qui stimule la production de FSH par l'hypophyse. En favorisant la croissance des follicules ovariens, la FSH stimule la production d'oestrogènes par eux, hormones qui atteignent un pic peu avant le milieu du cycle. L'augmentation du taux d'oestrogènes sanguins incite la production hypothalamique de LH-RH qui provoque une libération massive de LH hypophysaire.
phase ovulatoire:
Le taux de LH est maximal au milieu du cycle et provoque l'ovulation.
phase lutéale:
Après l'ovulation LH et FSH stimulent la formation du corps jaune (luteum), donc la production de progestérones et d'oestrogènes qui agissent sur l'utérus en le préparant pour la nidation éventuelle.
phase gestationnelle:
Si la nidation a lieu, le chorion produit une hormone, HCG (human chorionic gonadotropin), dont l'effet est analogue à celui de la LH: elle stimule la production de progestérones par le corps jaune et favorise le développement de l'embryon et du placenta. Le placenta produira lui-même des oestrogènes et progestérones durant la gestation.
phase menstruelle:
Si la nidation n'a pas lieu, le taux élevé de progestérones et d'oestrogènes circulantes exerce un effet négatif sur l'hypothalamus qui diminue alors sa synthèse d'hormones déclenchantes. Celles-ci ne sollicitant plus l'hypophyse, la production de FSH et LH diminue, et par conséquent la production d'hormones sexuelles par l'ovaire à la fin du cycle sexuel. Le corps jaune devient corps blanc. La paroi utérine privée de stimulation subit une vasoconstriction et une nécrose de ses couches superficielles, suivies d'une vasodilatation et d'une hémorragie.
Dans le contrôle de la gamétogenèse, l'hypothalamus représente l'échelon de la perception, l'antéhypophyse celui d'exécution et les gonades constituent les glandes cibles. La régulation de l'activité endocrine des gonades s'effectue suivant le principe du rétrocontrôle négatif: une diminution du taux circulant d'une hormone stéroïde sexuelle donnée signale la sécrétion hypothalamique de RH correspondante. L'hypophyse, sollicitée par la RH, élabore la stimuline correspondante. L'hormone hypophysaire active la production par la cible (gonade) de l'hormone sexuelle en quantité nécessaire pour rétablir le niveau "normal" de celle-ci. En concentration supérieure, cette hormone inhibe la production de RH hypothalamique. L'hypothalamus n'agit pas qu'en réponse à des concentrations d'hormones circulantes mais possède aussi une régulation cyclique fort complexe et peut répondre à des stimuli externes.
chez les amphibiens:
L'activité mitotique des ovogonies est saisonnière: les mitoses surviennent après chaque ponte et reconstituent un lot d'ovocytes qui, après croissance, formeront une ponte ultérieure; des nids d'ovogonies dormantes subsistent en permanence dans les ovaires. La croissance des follicules provenant d'une même vague mitotique saisonnière est synchrone. Le follicule mûr consiste en l'ovocyte entouré d'une membrane basale appelée enveloppe vitelline, d'une assise de cellules folliculaires sous-tendue d'une membrane basale et d'une thèque interne vascularisée. Celle-ci n'est pas immédiatement entourée par la thèque externe mais par un épithélium simple appelé épithélium ovarien interne, discontinu en une région qui est reliée à la thèque externe. Cette dernière englobe plusieurs follicules ovariens. Les follicules amphibiens subissent une croissance importante à la fin de leur cycle, ceci par l'accumulation de vitellus dans l'ovocyte pendant la fin de la phase diplotène (sous-phase de grand accroissement). L'ovocyte accroît son volume de 27 000 fois chez la grenouille. La sous-phase de petit accroissement correspond donc à la période de prévitellogenèse et celle de grand accroissement à celle de vitellogenèse.
Parallèles entre spermatogenèse et ovogenèse (voir schéma):
durée du cycle:
Le cycle se définit comme étant le temps nécessaire à la transformation d'une gonie nouvellement post-mitotique en gamète. La spermatogenèse est de plus courte durée que l'ovogenèse, différence d'autant plus marquée chez les Vertébrés supérieurs, chez qui un cycle spermatogénique ne dure que quelques semaines et un cycle ovogénique peut s'étendre sur toute la vie sexuelle de la femelle, des décennies parfois.
localisation temporelle de la période de multiplication:
Elle dure toute la vie génitale chez le mâle. Chez la femelle des anamniotes (poissons, amphibiens), des vagues de mitoses et de pontes alternent. Chez les amniotes, les ovogonies cessent de se multiplier pendant la vie foetale ou périnatale.
importance de l'accroissement:
Le volume des spermatocytes I augmente peu par rapport à celui des spermatogonies. L'accroissement des ovocytes I, qui accumulent des réserves parfois volumineuses, est considérable et de longue durée.
place de la méiose dans le cycle:
Sur la lignée mâle, après une phase de synthèse dans les spermatocytes I, les deux divisions méiotiques précèdent la différenciation morphologique en spermatozoïdes (spermiogenèse). La "différenciation" de l'ovocyte, qui consiste en réalité en accroissement volumétrique, a lieu dans la cellule diploïde.
résultat de la méiose:
À l'issue de la méiose, un spermatocyte I aura donné naissance à quatre spermatozoïdes identiques et fonctionnels. Un ovocyte subit des divisions méiotoques très inégales quant à la quantité de cytoplasme partagé entre les cellules-filles: une seule conserve toutes les réserves cytoplasmiques et sera fonctionnelle, l'ovocyte II puis l'ovotide, les globules polaires dégénérant.
Quelques caractéristiques des oeufs
Avant de décrire la fécondation, apportons quelques détails additionnels sur les caractéristiques des oeufs, outre ce qui a déjà été dit de la quantité de vitellus qu'ils renferment et de sa localisation, puisque ces caractéristiques influencent la fécondation et le développement embryonnaire.
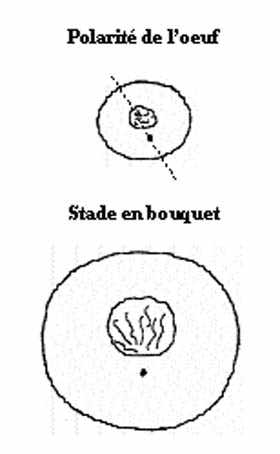
polarité:
À la fin de l'ovogenèse, on note une distribution inégale des composantes cellulaires dans le cytoplasme de l'oeuf, lui imputant une polarité. Dans les oeufs vitellins, le pôle animal se retrouve du côté du noyau (excentré) et le pôle végétatif du côté du vitellus. Lors des divisions méiotiques, le premier globule polaire est émis au pôle animal. Cette observation aide à définir le pôle animal dans les oeufs isolécithes. Outre la distribution préférentielle de certaines composantes cytoplasmiques, la polarité de l'oeuf mûr peut s'exprimer au niveau moléculaire. Par exemple, dans certains oeufs mammaliens, sans polarité évidente, la membrane cytoplasmique n'est pas identique sur toute sa surface. Durant l'ovogenèse, il apparaît une région membranaire dépourvue des microvillosités qui caractérisent le reste de la membrane. Les composantes moléculaires diffèrent quelque peu dans cette région lisse. De plus, le cytoplasme cortical sous la membrane lisse renferme plus de filaments d'actine, qui sont ancrés dans la membrane. Or, il s'avère que cette région membranaire lisse se situe à l'antipode du point d'émission du premier globule polaire (tiré de Talansky et al., Molec. Reprod. Devel., 28(1991):183-188). L'hétérogénéité de la membrane ovocytaire est probablement plus la règle que l'exception chez les différents groupes zoologiques. L'axe A-V de l'oeuf, déterminé durant l'ovogenèse, définit la future polarité antérieure - postérieure de l'embryon.
L'origine de la polarité est obscure. Elle peut être multiple. On soupçonne qu'elle est prédéterminé par la localisation cytoplasmique d'information dans l'oeuf. La position du centrosome (région cytoplasmique renfermant les centrioles) par rapport au noyau établit une polarité dont l'axe est la ligne passant par le noyau et le centrosome. Cette polarité affecte la distribution des composantes cytoplasmiques et même la structure du noyau. Au stade leptotène de la méiose, les chromosomes convergent vers la face du noyau la plus près du centrosome et présentent une configuration qui mérite à ce stade le nom de "stade en bouquet". Ces facteurs qui pourraient attribuer une polarité à l'oeuf sont intrinsèques à la cellule. Existe-t-il des facteurs extrinsèques ayant le même pouvoir? L'établissement de la polarité sera discuté au dernier chapitre.
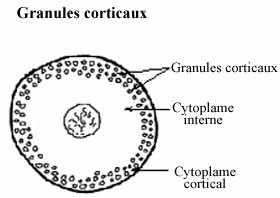
granules corticaux:
Vésicules cytoplasmiques sphériques renfermant des enzymes protéolytiques et retrouvées dans le cortex de l'oeuf sous la membrane cytoplasmique, les granules corticaux proviennent de l'appareil de Golgi dans le cytoplasme interne, durant la phase de grand accroissement. Ils jouent un rôle important lors de la fertilisation. Ils se retrouvent dans les oeufs de plusieurs groupes d'animaux: annélides, oursins, mollusques bivalves, poissons, anoures, certains mammifères (hamster, lapin, humain). Ils sont absents chez les insectes, les mollusques gastropodes, les urodèles, les reptiles, les oiseaux et certains mammifères (cobaye, rat).
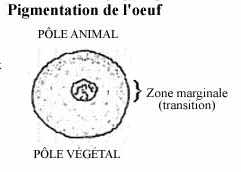
pigmentation:
Dans les oeufs pigmentés, les granules de pigment se retrouvent surtout dans le cytoplasme cortical. Le pigment peut être concentré au pôle animal, et particulièrement dans une moitié du pôle qui correspond à la future face dorsale de l'embryon. Le pigment peut être uniforme tout autour de l'oeuf et n'être que plus tard distribué inégalement aux cellules-filles, à la segmentation. Le pigment n'est probablement pas important pour le développement de l'embryon. Certains animaux ont des oeufs non pigmentés et se développent de la même façon que des espèces voisines pigmentées. Mais le pigment s'avère un repère permettant de suivre le développement embryonnaire.
Enveloppes:
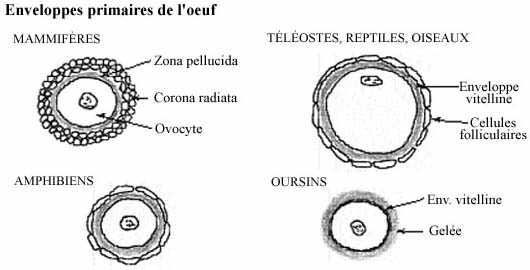
À part la membrane cytoplasmique qui délimite toute cellule, des enveloppes additionnelles entourent l'oeuf de toutes les classes zoologiques, sauf certaines éponges et certains coelentérés. Il existe deux catégories d'enveloppes: les enveloppes primaires se développent autour de l'oeuf quand il est encore dans l'ovaire tandis que les enveloppes secondaires sont sécrétées par les tissus des voies génitales parcourues par l'oeuf après l'ovulation.
enveloppes primaires (voir schéma):
La zona pellucida est sécrétée pas l'ovocyte (et les cellules folliculaires?) durant l'ovogenèse. Il s'agit d'une épaisse membrane basale, riche en mucopolysaccharides (PAS+) et en protéines fibreuses. Chez les groupes sous-mammaliens, l'on ne parle pas de zone pellucide mais de membrane ou enveloppe vitelline.
Les cellules folliculaires retenues autour de l'ovocyte et de sa zone pellucide (ou enveloppe vitelline) à l'ovulation ne font pas partie de l'enveloppe primaire. Elles seront détachées au fur et à mesure que l'oeuf parcourt l'oviducte.
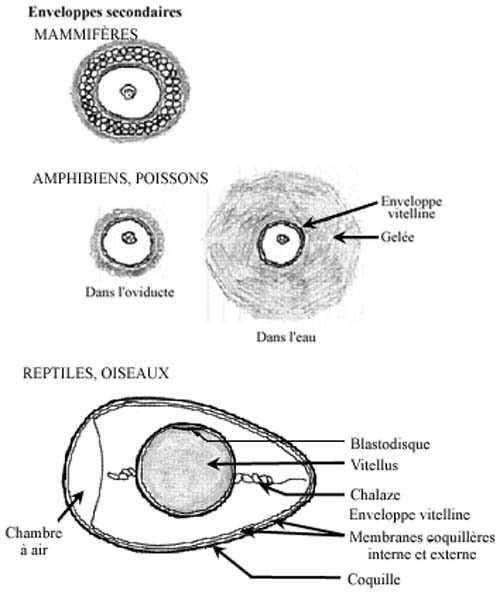
enveloppes secondaires (voir schémas):
Sécrétées par les cellules de la paroi de l'oviducte, les enveloppes secondaires sont déposées à la surface de l'oeuf. Chez les mammifères, une substance gélatineuse qui a la propriété d'absorber l'eau entoure la corona radiata. Chez les reptiles et les oiseaux, une couche fibreuse est ajoutée à l'enveloppe vitelline dès que l'oeuf atteint l'oviducte. Elle est considérée comme faisant partie de l'enveloppe vitelline. Dans les trois heures suivantes, l'albumen (blanc de l'oeuf) est déposé autour. Entourant l'albumen, les deux membranes coquillères, interne et externe, se composent de kératine. Finalement, la coquille de carbonate de calcium délimite l'oeuf entier. Nous reviendrons sur cela. Chez les amphibiens, les poissons et les invertébrés aquatiques, une gelée, ayant aussi la propension de retenir l'eau, permet aux oeufs de s'agglutiner, d'adhérer aux plantes aquatiques ou flottantes, aux roches, etc.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}