GASTRULATION (cours 5)
Le rythme effréné des divisions cellulaires, caractéristique de la segmentation, ralentit et l'embryon entre dans la phase de développement appelée gastrulation. En fin de segmentation, la blastula consiste en de nombreux blastomères dont les positions relatives ont été établies durant cette phase. À la gastrulation les cellules adopteront de nouvelles positions, donc de nouvelles voisines. Les mouvements coordonnés permettant la gastrulation, appelés mouvements morphogénétiques, impliquent l'embryon entier et aboutissent à la mise en place des 3 feuillets fondamentaux des métazoaires:
endoderme
mésoderme
ectoderme
Les coelentérés (hydres) et éponges ne développent pas de mésoderme.
Les mécanismes de gastrulation varient beaucoup dans le règne animal; on retrouve néanmoins une combinaison de quelques uns des mouvements morphogénétiques suivants:
embolie ou invagination:
Le repliement d'un feuillet cellulaire dans le blastocoele. Le mouvement d'embolie s'emploie surtout par les embryons dotés d'un blastocoele important et de blastomères végétatifs peu vitellins, de petite taille. Le feuillet cellulaire de l'hémisphère végétatif s'enfonce dans le blastocoele qui devient réduit. L'invagination délimite une structure tubulaire, l'archentéron, tube digestif embryonnaire, ouvert sur l'extérieur par le blastopore. On assiste ainsi à l'ébauche du tube digestif, qui a valu son nom au stade.
épibolie:
Lorsque les blastomères végétatifs sont trop volumineux pour s'invaginer dans le blastocoele ou que ce dernier est réduit ou absent, le feuillet cellulaire de l'hémisphère animal migre par dessus le feuillet végétatif et le recouvre. Ce dernier atteint donc une position interne de façon passive.
migration cellulaire ou ingression:
Migration de cellules individuelles, plutôt que d'un feuillet comme les mouvements précédents, dans le blastocoele où elles s'agencent pour former l'endoderme ou le mésoderme.
délamination:
Séparation d'un feuillet cellulaire pluristratifié en deux feuillets, l'un d'eux étant maintenant interne, dans le blastocoele.
prolifération polaire:
Multiplication active des blastomères du pôle végétatif vers l'intérieur du blastocoele. Contrairement à l'ingression, les cellules demeurent en masse compacte.
Des auteurs décrivent d'autres mouvements de gastrulation, en réalité des variations des cinq types décrits ici. Est-il nécessaire de leur attribuer un nom propre?
De façon similaire à la segmentation, la gastrulation se solde par une croissance totale minime. Toutefois, durant la gastrulation, les interactions nucléo-plasmiques sont plus importantes et plusieurs protéines sont synthétisées à partir du génome embryonnaire. De plus, le métabolisme devient oxydatif.
Exemples de gastrulation
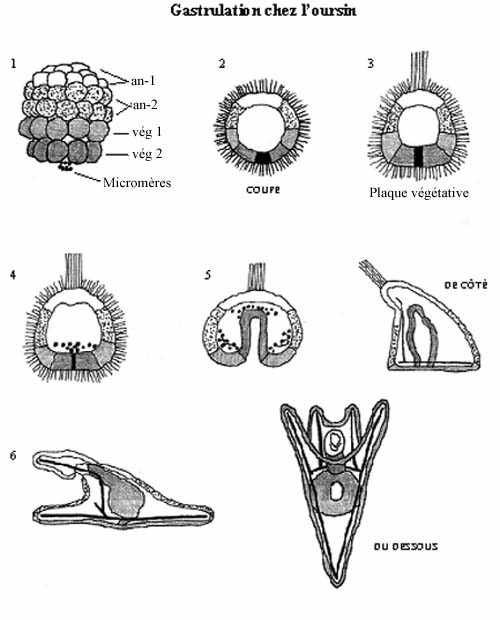
1) Oursin (échinoderme): oeuf oligolécithe; segmentation holoblastique, radiaire, totale et sub-égale (tailles cellulaires peu différentes).
La blastula d'oursin consiste en un feuillet cellulaire sphérique de 1000 à 2000 blastomères, selon les espèces, qui dérivent de différentes régions cytoplasmiques du zygote et qui ont des propriétés différentes. Elle peut être divisée en un hémisphère animal et un hémisphère végétatif.
Environ 24 heures après que la blastula se soit libérée de l'enveloppe de fertilisation (éclosion), la moitié la plus polaire de l'hémisphère végétatif s'aplatit; c'est la plaque végétative. Des micromères de la plaque végétative prolifèrent puis se détachent et migrent dans le blastocoele pour former le mésenchyme primaire, appartenant au feuillet intermédiaire: le mésoderme. Ces cellules fusionneront et formeront un syncitium, à l'origine du squelette de carbonate de calcium de la larve pluteus.
Le reste de la plaque végétative s'invagine dans le blastocoele. Ce faisant, les micromères restants se détachent, migrent dans le blastocoele et forment le mésenchyme secondaire, le reste du mésoderme. Ces cellules se dispersent à l'intérieur du blastocoele, où elles formeront les organes mésodermiques coelomiques. Ces organes présomptifs sont organisés en vésicules qui entreprendront un développement complexe.
En s'invaginant, la plaque végétative délimite une structure tubulaire, l'archentéron ou intestin embryonnaire, maintenant constitué que d'endoderme, qui s'ouvre à l'extérieur par un orifice appelé blastopore. Ce dernier marque la position du futur anus. Ce mouvement d'embolie de la plaque végétative semble se produire grâce à des forces intrinsèques à la plaque, et non pas parce que les cellules adjacentes poussent sur elle pour la forcer à s'invaginer.
L'hémisphère animal et la première rangée de macromères végétatifs (vég-1) forment ensemble l'ectoderme, qui recouvre les deux autres feuillets complètement internalisés.
À un endroit précis, l'ectoderme s'invagine légèrement et se rapproche du toit de l'archentéron. Les deux entrent éventuellement en contact et fusionnent, formant la membrane stomodéale, qui se perfore: le stomodeum, future bouche de l'animal. Le tube digestif continu, de l'orifice d'entrée à l'orifice de sortie, est ainsi formé. Chez les deutérostomes, l'anus est donc formé avant la bouche.
La mise en place des 3 feuillets: ectoderme, mésoderme et endoderme, est achevée et ceux-ci sont emboîtés, avec l'endoderme le plus interne et l'ectoderme externe.
La gastrulation de l'oursin combine les mouvements morphogénétiques suivants:
1) prolifération polaire de micromères
2) leur migration dans le blastocoele, formant le mésenchyme primaire
3) invagination (embolie) indépendante de la plaque végétative, formant l'archentéron
4) migration des autres micromères de l'archentéron, formant le mésenchyme secondaire
5) invagination d'une portion de l'ectoderme
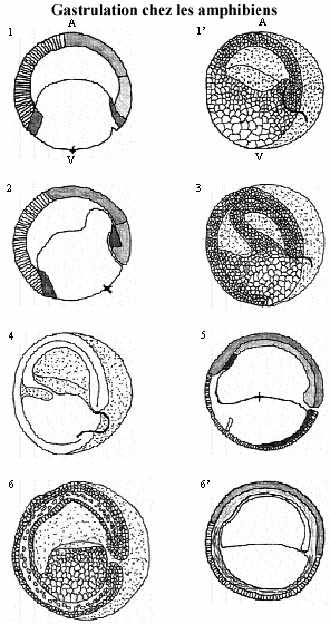
2) Amphibiens: oeuf mésolécithe hétérolécithe: vitellus ségrégé du cytoplasme actif après la fécondation, noyau au pôle animal, donc polarité évidente. Chez plusieurs espèces, le cytoplasme cortical, plus visqueux que l'interne, renferme des pigments de mélanine, surtout dans l'hémisphère animal. Les produits accumulés pendant l'ovogenèse se retrouvent dans le cytoplasme interne, selon un gradient décroissant du pôle animal au pôle végétatif, sauf pour les réserves vitellines qui suivent un gradient inverse. La segmentation est holoblastique, radiaire et inégale. La blastula renferme un blastocoele réduit.
La segmentation rapide n'est synchrone que pendant les 12 premières divisions. Le rythme des divisions ralentit et perd de sa synchronie, certaines cellules demeurant plus longtemps en interphase. C'est alors que dans ces cellules commencent les premières synthèses d'ARN. De même, des changements dans la motilité cellulaire s'amorcent et annoncent les premiers signes de gastrulation.
La gastrulation commence sous l'équateur, à un point correspondant à l'antipode du point d'entrée du spermatozoïde. Chez les espèces pigmentées, ce point de départ de la gastrulation se localise dans la région du croissant gris.
Le croissant gris résulte de la réorganisation du cytoplasme de l'oeuf suite à l'entrée du spermatozoïde. La portion de cytoplasme à 180o de l'entrée du spermatozoïde décrit la forme d'un croissant, gris parce que pigmenté. Ce point marque aussi la future face dorsale de l'embryon. Le point d'entrée du spermatozoïde marque donc la future face ventrale. L'axe dorso-ventral est ainsi établi, ainsi que l'axe de symétrie bilatérale, qui coupe ces deux faces. Tout au cours de la segmentation le croissant gris s'estompe.
Donc au point du croissant gris, le feuillet de blastomères s'invagine légèrement, invagination prenant la forme d'un sillon horizontal à la surface de l'oeuf: c'est la lèvre dorsale du blastopore (LDB). Au fur et à mesure des mouvements suivants, des lèvres latérales (2) et ventrale (1) se dessineront; il en résultera la formation d'une ouverture ronde, le blastopore.
Alors que s'invagine légèrement le feuillet de blastomères du croissant gris, le feuillet de l'hémisphère animal entreprend un mouvement d'épibolie qui converge vers le blastopore. Quand ces blastomères atteignent la LDB, ils poursuivent leur migration à l'intérieur du blastocoele en glissant sous l'hémisphère animal. Ce faisant, ils repoussent de plus en plus vers l'intérieur les cellules qui formaient initialement la LDB. Ainsi, la LDB est composée de cellules continuellement en changement, car successivement repoussées vers l'intérieur.
Ainsi, les blastomères du plancher du blastocoele qui, à cause de leur taille, étaient incapables de s'invaginer activement, sont maintenant tirés dans le blastocoele par le mouvement d'épibolie des blastomères animaux. Ils sont donc passivement amenés à l'intérieur du blastocoele.
Le front cellulaire d'épibolie progresse sur toute la surface interne du blastocoele et délimite l'archentéron, structure tubulaire à l'intérieur du blastocoele. Comme de plus en plus de cellules pénètrent à l'intérieur du blastocoele, augmentant la taille de l'archentéron, le blastocoele devient graduellement réduit et repoussé à l'antipode du blastopore. L'archentéron se trouve ainsi délimité, dorsalement, par la portion du feuillet animal qui a été internalisée (activement) et, ventralement, par les blastomères végétatifs qui ont été internalisés (passivement).
Le mouvement d'épibolie se poursuit au point que l'archentéron devient oblitéré par le front de cellules, qui forme le bouchon vitellin bouchant le blastopore. Un mouvement d'épibolie des cellules externes les fait recouvrir le bouchon vitellin. Le blastopore n'a plus que l'aspect d'une fente verticale, le futur anus, oblitéré pour le moment. Toutes les cellules qui restent en position externe constituent l'ectoderme.
Les toutes premières cellules à former la LDB, constituant le front de migration, sont des blastomères végétatifs (initialement situés juste sous l'équateur); amenées à l'intérieur de l'embryon, elles formeront une portion de l'épithélium du tube digestif, donc de l'endoderme. Les blastomères suivants, provenant de l'hémisphère animal juste sus l'équateur, sont dits mésoblastiques, précurseurs du mésoderme de la tête. Les prochains blastomères à s'internaliser par la LDB, dits chordamésodermiques, sont à l'origine de la notochorde, structure non retrouvée chez les invertébrés. Les blastomères suivants formeront le reste du mésoderme. Les gros blastomères du plancher de l'archentéron complètent l'épithélium de l'archentéron. Donc pendant un certain temps, le toit de l'archentéron est formé d'un feuillet mésodermique.
Un espace intercellulaire sépare le feuillet ectodermique du feuillet mésodermique dans l'hémisphère animal et du feuillet endo-mésodermique dans l'hémisphère végétatif. Les cellules mésodermiques se détachent et migrent, plus ou moins individuellement, latéralement et ventralement dans cet espace. Ce faisant, l'endoderme referme le toit de l'archentéron. Celui-ci devient donc délimité complètement par l'endoderme. À la fin de la gastrulation, l'embryon comprend les 3 feuillets fondamentaux emboîtés.
La gastrulation amphibienne est donc marquée principalement par des mouvements d'invagination, d'épibolie et de migration. À 18°C elle se produit en 24 heures environ.
Contrairement à l'oursin, chez qui la gastrulation commence au pôle végétatif, la gastrulation amphibienne commence près de l'équateur de la blastula, où les cellules contiennent moins de vitellus que celles du pôle végétatif, quoiqu'elles en contiennent suffisamment pour ne pouvoir s'invaginer significativement.
La LDB marque véritablement le point de départ de la gastrulation. En effet, si la LDB est transplantée dans une autre région de la blastula, la gastrulation pourrait s'amorcer à cet endroit.
Puisque la LDB occupe dans la blastula l'endroit correspondant au croissant gris de l'oeuf fécondé, on a longtemps soupçonné que cette région contenait des facteurs cytoplasmiques capables d'initier la gastrulation. La localisation du blastopore est déterminée par un ou des facteurs dans le cytoplasme interne, plutôt que cortical, de l'oeuf. Les réarrangements du cytoplasme occasionnés par l'entrée du spermatozoïde sont responsables de la distribution asymétrique de facteurs cellulaires. Le facteur initiateur de la gastrulation ne se trouve pas dans les cellules pigmentées du croissant gris mais dans des cellules végétatives profondes. Le facteur en question n'est exprimé qu'à partir d'un certain stade de la segmentation. Les cellules végétatives profondes le contenant induisent les cellules superficielles du croissant gris à initier la gastrulation et à former la face dorsale du corps. Un candidat au titre de facteur initiateur de la gastrulation est l'activine: un facteur de croissance peptidique.
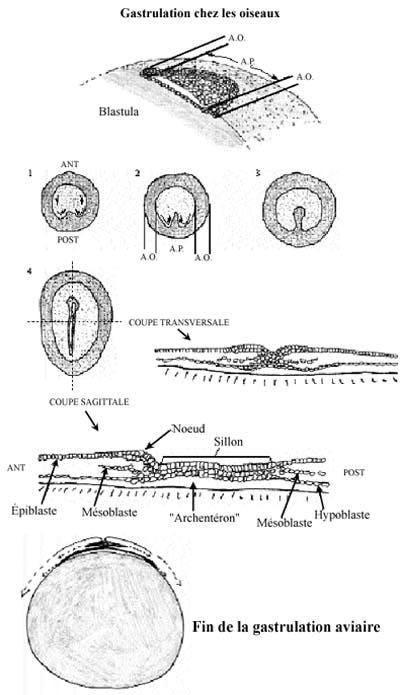
3) Oiseaux: oeuf télolécithe; segmentation partielle, n'affectant que le disque germinal et résultant en la formation d'une discoblastula sur une énorme masse de vitellus. Par délamination, des blastomères du disque germinal migrent dans la cavité sous-germinale et forment l'hypoblaste, les cellules de surface formant l'épiblaste. Entre l'épiblaste et l'hypoblaste se retrouve le blastocoele.
Chez le poulet, la gastrulation commence dès les premières heures d'incubation. Le premier indice en est l'apparition d'un épaississement dans la région postérieure de l'aire pellucide, qui progresse antérieurement sur la ligne médiane, s'étendant sur 50 à 75% de l'aire pellucide. Cet épaississement témoigne d'une migration convergente des cellules de l'aire pellucide vers le centre postérieur puis vers l'avant, et constitue la ligne primitive, terminée par un renflement antérieur, le noeud primitif ou de Hensen. La ligne primitive marque l'axe antérieur-postérieur de l'embryon. Elle constitue la caractéristique structurale majeure de la gastrulation des oiseaux, comme de celle des reptiles et des mammifères (des amniotes).
Comme les cellules de l'aire pellucide convergent pour tracer la ligne primitive, une dépression se creuse dans celle-ci, le sillon primitif. Le sillon primitif sert de blastopore au travers lequel les cellules de l'épiblaste migrent dans le blastocoele. Le sillon primitif des oiseaux, reptiles et mammifères est donc analogue au blastopore amphibien. Comme pour ce dernier, le sillon primitif consiste en une population cellulaire continuellement en changement. Au centre du noeud primitif se creuse pareillement une dépression en forme d'entonnoir et au travers laquelle les cellules de l'épiblaste migrent dans le blastocoele. Le noeud primitif est analogue à la LDB des amphibiens.
Les premiers blastomères de l'épiblaste à migrer dans le blastocoele empruntent le sillon primitif, migrent antérieurement, déplaçant les cellules hypoblastiques de la région antérieure de l'embryon, et formeront une portion du tube digestif, endodermique. Une partie de l'endoderme est maintenant amenée à l'intérieur de l'embryon.
Les prochains blastomères à pénétrer dans le blastocoele le font par le noeud primitif et migrent antérieurement, en demeurant entre l'endoderme et l'épiblaste; ils formeront le mésoderme de la tête et le chordamésoderme, lui aussi maintenant internalisé. La migration antérieure de toutes ces cellules a soulevé la région antérieure de l'épiblaste et résulte en la formation du bourgeon de la tête. Le blastodisque s'allonge.
Les dernières cellules migrant au travers du sillon primitif se séparent en deux courants une fois dans le blastocoele. Un courant migre profondément et déplaçe l'hypoblaste latéralement et postérieurement. Les cellules de ce courant de migration donneront naissance au reste des structures endodermiques embryonnaires et extra-embryonnaires. Les cellules de l'autre courant s'étendent en un feuillet lâche, entre l'endoderme et l'épiblaste et sont à l'origine du reste du mésoderme embryonnaire et extra-embryonnaire.
À 22 heures d'incubation, la majorité des cellules endodermiques présomptives se trouvent à l'intérieur de l'embryon, et des cellules mésodermiques présomptives continuent de migrer pour une période de temps prolongée.
Une seconde phase de la gastrulation s'entame: le sillon primitif régresse petit à petit caudalement, le noeud primitif occupant donc une position de plus en plus postérieure. C'est ainsi que se forme la notochorde, selon un gradient antéro-postérieur, par les dernières cellules mésodermiques à être mises en place. Finalement, en position complètement postérieure, le noeud primitif forme la région anale présomptive, de la façon deutérostome. Ce qui reste de l'épiblaste après ces migrations cellulaires, les cellules de surface, constitue l'ectoderme.
Contrairement à ce observé chez les amphibiens, chez qui des feuillets cellulaires migrent dans le blastocoele, chez les oiseaux ce sont des cellules individuelles qui migrent. À l'intérieur du blastocoele, plutôt que de créer un feuillet dense, elles forment un tissu lâche, particulièrement le mésoderme. Les cellules migrantes se déplacent par la formation de pseudopodes sur un substrat de polysaccharides, notamment l'acide hyaluronique, fabriqué par les cellules épiblastiques dès le début de la gastrulation. Si l'enzyme hyaluronidase est injectée dans le blastocoele, les cellules s'agrègent et ne migrent pas ou peu. L'acide hyaluronique est donc requis pour garder les cellules migrantes séparées les unes des autres.
Chez les oiseaux, le feuillet endodermique ne compose pas de structure tubulaire, comme l'archentéron de l'oursin ou de la grenouille, mais un feuillet plat. Il ne se forme donc pas de véritable archentéron dans la gastrula des oiseaux. Les trois feuillets ne sont pas emboîtés mais superposés.
Une conséquence de la gastrulation en deux étapes des amniotes est l'émergence d'un gradient antéro-postérieur de développement. Tandis que la portion postérieure de l'embryon est encore en gastrulation, la portion antérieure amorce l'étape subséquente de développement, l'organogenèse. La région antérieure de l'embryon sera longtemps plus avancée dans son développement que la région postérieure.
Pendant que se poursuit la gastrulation, des cellules ectodermiques, reliées par des jonctions serrées, entament un mouvement d'épibolie qui les fera entourer le vitellus. Leur surface externe adhère à l'enveloppe de fertilisation et les cellules s'étendent contre elle. Ceci prend 4 jours à s'accomplir et requiert la formation de nouvelles cellules, par mitoses. Finalement, l'endoderme remplace l'hypoblaste autour du vitellus, l'ectoderme entoure incomplètement le vitellus, et le mésoderme extra-embryonnaire se place entre les deux. Le vitellus ainsi entouré des trois feuillets prend le nom de sac vitellin. Les trois feuillets extra-embryonnaires du sac vitellin sont continus avec les trois feuillets embryonnaires. Nous y reviendrons.
Chez les oiseaux, c'est l'hypoblaste de la blastula qui induit la formation de la ligne primitive; l'enlever retarde tout développement subséquent, tant que l'épiblaste n'a pas généré, par délamination, une autre couche hypoblastique. L'hypoblaste dirige la formation et la directionnalité de la ligne primitive.
4) Mammifères: oeuf alécithe; segmentation holoblastique rotationnelle.
Bien que les mouvements morphogénétiques de la gastrulation des reptiles et oiseaux aient évolué en adaptation à des oeufs riches en vitellus, ils ont été retenus chez les mammifères.
L'épiblaste embryonnaire de la blastula mammalienne est similaire à l'épiblaste aviaire. À son bout postérieur un épaississement produira la ligne primitive et le noeud primitif, qui se creuseront en sillon primitif et gouttière primitive au travers lesquels migreront les précurseurs endodermiques et mésodermiques. Ce processus est semblable à celui retrouvé chez les oiseaux.
Axes et polarité:
Revenons brièvement à ces notions, nous limitant aux Vertébrés. On réalise que le premier axe à se manifester est l'axe dorso-ventral. Cet axe se trouve à définir quelle est la face dorsale de l'embryon et quelle est sa face ventrale. Une ligne coupant cet axe sépare l'embryon en 2 moitiés, gauche-droite, et établit la symétrie bilatérale. Toutefois, quand on examine le développement rétrospectivement, on réalise que l'axe dorso-ventral, la
symétrie bilatérale, était mis en évidence plus tôt; il a été déterminé plus tôt qu'il n'apparaît. En effet, le point d'entrée du spermatozoïde a amené un réaménagement cytoplasmique important. Ce point correspond au pôle ventral de l'embryon et celui diamétralement opposé au pôle dorsal. Cependant, le spermatozoïde agissait sur un cytoplasme déjà polarisé, qui imposait des contraintes à son entrée, et la polarité de l'oeuf est ultimement à l'origine de cet axe. Le pôle animal sera la face dorsale de l'embryon et le pôle végétatif la face ventrale. Nous avons vu que dans les oeufs non vitellins d'autres indices dévoilent une polarité intrinsèque. Les bases moléculaires et génétiques de cette polarité font actuellement l'objet de nombreuses investigations.
En considérant le développement rétrospectivement, l'axe antéro-postérieur se manifeste dès le début de la gastrulation. Le point de départ de la gastrulation: blastopore ou noeud primitif, correspond à l'emplacement du futur anus (le pôle postérieur); l'autre bout de l'embryon est inévitablement la future tête. Ici aussi la polarité a été déterminée plus tôt qu'elle n'apparaît. On le sait particulièrement des amphibiens chez qui la localisation du croissant gris, apparaissant à la fécondation, signale le point de départ de la gastrulation. Les bases moléculaires et génétiques de cette polarité font également l'objet de recherches intensives. Elle implique probablement un gradient de molécules cytoplasmiques dans l'oeuf, avant ou après la fécondation.
Il faut réaliser que les deux polarités sont dissociables. L'existence d'un axe, que ce soit le dorso-ventral ou l'antéro-postérieur, n'amène pas celle de l'autre.
Une autre forme de polarité ou d'axe existe; il s'agit de la latéralisation. L'axe dorso-ventral, ou la symétrie bilatérale, a amené la création de 2 moitiés dont on ne sait toutefois pas encore laquelle est la gauche et laquelle la droite tant que l'axe antéro-postérieur n'est pas établi. Ceci accompli, les moitiés gauche et droite sont en principe identiques, sauf qu'une est l'image miroir de l'autre. Cependant, on sait pertinemment, avant même d'étudier l'organogenèse, que tout dans le vertébré n'est pas pair. Qu'est-ce qui détermine que des organes uniques se localisent à gauche ou à droite? Il semble que la matrice extracellulaire joue un rôle dans cette détermination.
Territoires présomptifs:
Différentes techniques permettent de marquer une cellule ou un groupe de cellules et d'en suivre l'évolution temporelle. Ceci a permis de décrire les mouvements de gastrulation et de tracer le devenir des cellules de la gastrula, établissant ainsi la carte des territoires présomptifs de l'embryon.
Les Métazoaires ont évolué sur plusieurs lignées. La majorité peuvent être groupés dans deux embranchements. Les deutérostomes subissent une segmentation radiaire, les blastomères sont à 90o les uns aux autres et possèdent une capacité de régulation (dernier chapitre), et à la gastrulation l'anus est formé avant la bouche. Les protostomes ont une segmentation spiralée, les blastomères sont à angle aigu les uns aux autres et possèdent moins de capacité de régulation, et la bouche se forme avant l'anus.
{kind=link}
{kind=link}
{kind=link}