NUTRITION DE L'EMBRYON ET STRUCTURES EXTRA-EMBRYONNAIRES (cours 11)
L'embryon en formation est un organisme qui requiert O2 et substances nutritives pour se maintenir et se développer. Cependant, il ne possède pas encore les organes capables d'accomplir les fonctions de respiration, nutrition et excrétion. L'apport et l'utilisation d'O2 et de nourriture, ainsi que l'élimination du CO2 et des déchets métaboliques doivent donc être organisés de façon spéciale et spécifique à la vie embryonnaire.
L'embryon peut avoir recours à deux types de ressources:
- les réserves de l'oeuf, emmaganisées pendant l'ovogenèse. Leur nature et quantité dépendent du type d'oeuf à partir duquel se développe l'embryon.
- les substances exogènes, extraites de l'environnement. Leur nature et quantité dépendent de l'environnement dans lequel se développe l'embryon.
En relation avec l'apport des substances nécessaires à l'embryon, les animaux ont développé différentes stratégies de développement: oviparité, ovoviviparité et viviparité.
1) Oviparité:
L'oviparité est le mode de développement selon lequel tout ou presque tout le développement de l'embryon se produit dans l'oeuf après qu'il ait été pondu. C'est le cas de tous les animaux à fécondation externe, chez qui tout le développement a lieu après la ponte, et de plusieurs animaux à fécondation interne qui pondent leurs oeufs peu après la fécondation.
Parmi les animaux ovipares, ceux qui se développent à partir d'un oeuf pauvre en réserves vitellines atteignent tôt un stade leur permettant de se nourrir et de se maintenir individuellement. Par exemple les échinodermes développent un stade larvaire dans les 35 à 40 heures suivant la fécondation.
Les embryons (et les larves) se développant en milieu aquatique absorbent beaucoup d'eau, qui constitue d'ailleurs un fort pourcentage de leur composition. En milieu marin, ils puisent en plus les sels minéraux nécessaires à édifier leur squelette (ex. le squelette calcaire de plutéus); ceux se développant en eau douce doivent avoir recours aux réserves cytoplasmiques pour les sels minéraux. Les lipides et le glycogène utilisés comme source d'énergie proviennent de réserves cytoplasmiques. Plus celles-ci sont faibles plus le stade larvaire est précoce.
Attention! toutes les larves ne sont pas capables de se nourrir. Il y a des espèces sédentaires qui développent des stades larvaires à cause de leur mobilité, permettant la dispersion de l'espèce.
La ponte précoce de l'oeuf présente une difficulté nouvelle pour les animaux terrestres, à fécondation interne. Les amphibiens et certains reptiles (certaines tortues) ont fait un compromis, ils retournent à l'eau pour la ponte et c'est dans le milieu aquatique que se déroule le développement embryonnaire. Ils se nourrissent à partir de leur réserves vitellines et développent une circulation vitelline, et la respiration des larves d'amphibiens est branchiale
Les autres reptiles et les oiseaux, quant à eux, ont développé des membranes imperméables qui empêchent l'absorption d'eau ou sa perte par évaporation. Ils se développent à partir d'un oeuf riche en réserves vitellines. Seul l'O2, nécessaire aux réactions oxydatives, est puisé de l'extérieur; le CO2 est pareillement expulsé. À part le transport des gaz, l'oeuf constitue un système clos. On dit que c'est un oeuf cléidoïque, comme une boîte. L'embryon se développe entièrement aux dépens des réserves.
De façon indépendante des reptiles et oiseaux, des arthropodes, dont les insectes terrestres, ont également développé des oeufs cléidoïques.
L'accumulation de réserves dans l'oeuf, tant chez les animaux aquatiques qui ne développent pas de stades larvaires, comme les mollusques céphalopodes et les poissons élasmobranches, que chez les animaux terrestres, comme les reptiles et oiseaux, permet à l'animal d'atteindre un stade de développement plus avancé avant la naissance (l'éclosion) que les espèces développant un stade larvaire; ceci augmente leurs chances de survie.
- oviparité chez les oiseaux:
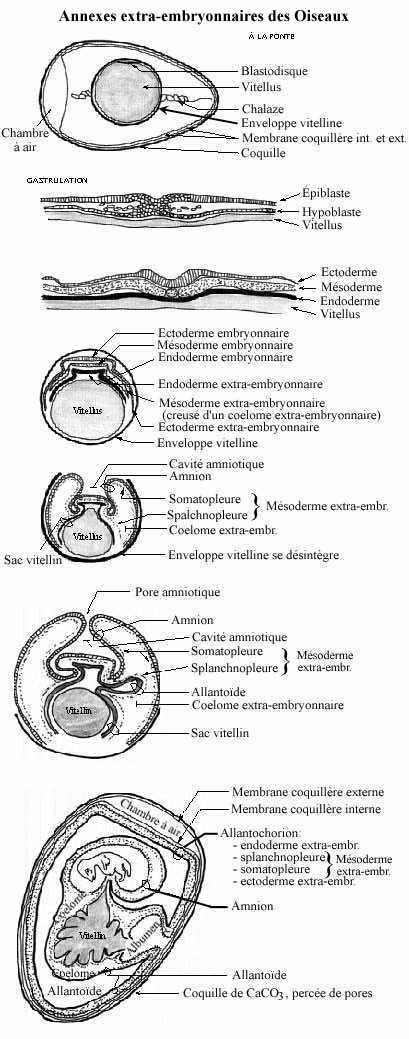
À l'ovulation, l'oeuf descend dans l'oviducte où a lieu la fécondation. L'enveloppe de fécondation se forme peu après, puis une portion de l'albumen. En 3 heures environ, l'oeuf fécondé, en début de segmentation, atteint la portion inférieure de l'oviducte (utérus). Le reste de l'albumen, les membranes coquillères et la coquille se forment dans les 20-24 heures suivantes. L'embryon avec le vitellus sous-jacent subit des rotations à l'intérieur de sa coquille, dont témoignent d'ailleurs les chalazes, rotations qui indiquent l'axe antéro-postérieur de l'embryon. Finalement, la ponte a lieu; l'embryon d'oiseau se trouve au stade de blastodisque, prêt à entamer la gastrulation. L'embryon même et le vitellus sont entourés de l'enveloppe de fécondation, de l'albumen, des deux membranes coquillères et de la coquille.
L'albumen (le "blanc") se compose à 85% d'eau et à 15% de protéines, dont 94% d'albumine. Les chalazes consistent en deux régions particulièrement denses de l'albumen rattachées à la membrane coquillère interne; elles maintiennent l'embryon en place, au centre de la coquille.
Les membranes coquillères se composent de kératine. Les deux sont intimement apposées, sauf à une extrémité où l'interne adhère à l'albumen et l'externe à la coquille. Entre les deux: la chambre à air. La coquille consiste surtout en carbonate de calcium (CaCO3). Elle est percée de nombreux pores remplis d'une protéine semblable à du collagène. Les pores permettent les échanges d'O2 et de CO2. Dans l'oeuf de poule, par exemple, la coquille possède environ 7 000 pores de 40-50mm de diamètre.
Suivant la gastrulation et en même temps que disparaît graduellement l'enveloppe de fécondation, un mouvement d'épibolie entraîne les trois feuillets: endoderme, mésoderme et ectoderme, à entourer le vitellus, bien qu'incomplètement. Ce mouvement permet la distinction entre feuillets embryonnaires et extra-embryonnaires. L'embryon se soulève du sac vitellin, lui étant rattaché par la tige vitelline. Ce faisant, le mésoderme extra-embryonnaire, comme le mésoderme embryonnaire, se délamine en deux feuillets par une cavité, le coelome extra-embryonnaire. Le feuillet accolé à l'ectoderme est appelé somatopleure et celui accolé à l'endoderme splanchnopleure. Ensemble, l'ectoderme et la somatopleure forment le chorion. Pour certains auteurs, la somatopleure comprend et l'ectoderme et la couche mésodermique somatique (pariétale), de même que la splanchnopleure comprend et l'endoderme et la couche splanchnique (viscérale) du mésoderme. Nous n'en faisons pas ainsi dans ce cours.
Une seconde épibolie élève l'ectoderme extra-embryonnaire et le feuillet somatique du mésoderme extra-embryonnaire par dessus l'embryon, formant ainsi l'amnion. Tout d'abord ouvert vers l'extérieur (l'albumen) par le pore amniotique, l'amnion vient à se refermer autour de l'embryon. Il est rempli du liquide amniotique qui provient de la déshydratation de l'albumen. L'embryon baigne donc dans ce liquide.
Les mouvements d'épibolie font que les deux feuillets du mésoderme extra-embryonnaire sont incorporés à des structures différentes: la somatopleure se trouve incorporée à l'amnion et la splanchnopleure au sac vitellin et à l'allantoïde (voir plus loin).
Des vaisseaux sanguins se différencient in situ dans le mésoderme splanchnique et vascularisent le sac vitellin. Des enzymes sécrétées par l'endoderme extra-embryonnaire dégradent le vitellus, ses produits passant à la circulation vitelline et à la circulation embryonnaire (revoir la chapitre précédent) pour nourrir l'embryon.
À la jonction du sac vitellin et de l'intestin postérieur se dessine un diverticule de l'endoderme qui entraîne avec lui le mésoderme splanchnique; il s'agit de l'allantoïde (allantoine). L'allantoïde étant continu avec le cloaque, il en reçoit les contenus (digestifs, urinaires et génitaux). L'allantoïde envahira rapidement tout le coelome extra-embryonnaire jusqu'à entourer l'amnion et le sac vitellin. L'albumen, de plus en plus utilisé, rapetisse. En occupant tout le coelome extra-embryonnaire, l'allantoïde devient délimité par 4 couches: endoderme extra-embryonnaire, splanchnopleure, somatopleure et ectoderme extra-embryonnaire; on parle alors d'allantochorion. Des vaisseaux sanguins se développent dans le mésoderme, connectés par la tige vitelline à l'aorte descendante (pour les artères allantoïdiennes) et à la veine cave inférieure (pour les veines allantoïdiennes).
L'embryon aviaire jouit donc de 3 annexes: le sac vitellin, l'amnion et l'allantoïde.
- Le sac vitellin renferme les réserves vitellines, sa paroi vascularisée comprend endoderme extra-embryonnaire et splanchnopleure, et il est relié à l'embryon par la tige vitelline qui véhicule les vaisseaux vitellins.
- L'amnion, avec une paroi d'ectoderme extra-embryonnaire et de somatopleure, délimite la cavité amniotique remplie du fluide amniotique dans lequel baigne l'embryon; il isole l'embryon, le protège et sert en même temps d'amortisseur.
- L'allantoïde, plutôt l'allantochorion, sert de réservoir des déchets éliminés par l'embryon et sa paroi vascularisée constitue le site d'échanges respiratoires (via les pores coquillères). De plus, des sels de la coquille peuvent être absorbés par l'allantochorion et transférés à l'embryon pour l'édification de son squelette.
À l'éclosion, amnion et allantoïde sont éliminés avec la coquille, l'albumen a été utilisé et le vitellus restant se rétracte dans l'abdomen de l'animal.
2) Ovoviviparité:
L'ovoviviparité est le mode de développement selon lequel l'embryon se développe à l'intérieur de l'organisme maternel, retenus dans les oviductes, jusqu'à un stade plus ou moins avancé selon les espèces, sans que la mère ne lui pourvoie de substances nutritives. Cette stratégie permet d'augmenter davantage les chances de survie des petits. Chez certains, l'éclosion se produit peu après la ponte des embryons tandis que chez d'autres, telles certaines salamandres, l'éclosion s'effectue dans l'organisme maternel.
L'ovoviviparité typique suppose que l'embryon est entièrement nourri à partir de réserves de l'oeuf, mais il y en a qui absorbent des substances du fluide utérin. Le degré de dépendance de l'embryon de l'une ou l'autre source: réserves de l'oeuf ou fluide utérin, varie d'une extrême à l'autre. Les mammifères marsupiaux ne possèdent pas de placenta et se développent grandement aux dépens du fluide utérin maternel. Leur naissance a lieu précocement, comparativement aux mammifères euthériens (placentaires).
3) Viviparité:
La viviparité est le mode de développement selon lequel l'embryon se développe à l'intérieur de la mère et établit une connexion directe avec elle. Les substances nutritives passent de la mère à l'embryon. La connexion s'établit grâce à un organe spécial, le placenta. Le placenta comprend une excroissance de l'embryon attachée à une portion de l'utérus maternel modifiée à cet effet. L'apport de substances nutritives à l'embryon s'effectue de façon beaucoup plus efficace par le placenta que par le fluide utérin. Le placenta permet un plus grand accroissement de l'embryon avant la naissance et augmente considérablement ses chances de survie.
Viviparité et placenta se sont développés indépendamment chez différents groupes zoologiques: tuniciés, poissons élasmobranches, mammifères. Le mode d'origine et la structure du placenta diffèrent chez ces groupes.
- viviparité chez les mammifères placentaires:
L'une des premières différenciations de l'embryon des mammifères est celle des cellules qui formeront le placenta, comme nous l'avons déjà abordé dans le cadre de la segmentation.
L'implantation (nidation) de l'embryon mammifère dans l'utérus maternel s'accomplit au stade blastocyste. Une prolifération du trophoblaste au pôle animal se caractérise par des caryocinèses plus nombreuses que les cytocinèses; il en résulte un syncitiotrophoblaste. Le reste du trophectoderme se compose de cellules complètes: le cytotrophoblaste. C'est le syncitiotrophoblaste qui envahit la paroi utérine, préalablement préparée par l'action d'hormones: FSH, progestérone. Au site d'implantation, la vascularisation s'active dans la paroi utérine (multiplication de vaisseaux existants, formation de vaisseaux de novo ).
C'est alors que l'embryon entreprend la gastrulation. Le mésoderme embryonnaire se continue (par épibolie) en mésoderme extra-embryonnaire qui se délamine. La couche somatique (pariétale) double l'ectoderme extra-embryonnaire pour former ensemble l'amnion qui enveloppera l'embryon. La couche splanchnique (viscérale) se vascularise et double l'endoderme extra-embryonnaire du sac vitellin et de l'allantoïde, et elle double le cytotrophoblaste, les deux formant le chorion. Le sac vitellin et l'allantoïde ont très peu d'importance chez les mammifères placentaires.
Le chorion et la portion de la paroi utérine, vascularisée, à laquelle il est fusionné constituent ensemble le placenta. Le sac vitellin et l'allantoïde réduits, comprenant le mésoderme viscéral extra-embryonnaire vascularisé, forment ensemble le cordon ombilical, qui relie l'embryon au placenta. Dans le placenta, les vaisseaux utérins et embryonnaires (splanchnopleure) s'apposent pour permettre les échanges respiratoires et métaboliques, les deux étant séparés par le mince cytotrophoblaste qui agit comme barrière contre les substances qui ne devraient pas atteindre la circulation embryonnaire.
Chez les mammifères placentaires, c'est la cavité amniotique qui envahit le coelome extra-embryonnaire, et non l'allantoïde comme chez les oiseaux. L'amnion s'accollera au chorion.
Selon l'importance du contact entre l'embryon et l'utérus maternel, on décrit deux types principaux de placenta:
- le placenta de contact: les tissus embryonnaire et maternel s'accollent mais la paroi utérine subit relativement peu de modifications; elle demeure quasiment intacte après l'implantation. À la naissance, l'expulsion du placenta se fait sans hémorragie. C'est le type de placenta retrouvé chez le porc, le cheval et la vache.
- le placenta décidu: la pénétration du trophoblaste dans la paroi utérine la modifie considérablement; il y a contact intime entre les tissus embryonnaire et maternel et la paroi utérine se referme complètement autour de l'embryon. Le chorion forme de nombreuses villosités qui augmentent la surface d'échange vasculaire entre les deux organismes. À la naissance, l'expulsion du placenta oblige une portion de la paroi utérine à être rejetée, détachée du reste de l'utérus; il s'ensuit une hémorragie. Ce type de placenta se rencontre chez le chien, le rat et l'humain.
En plus du transfert de gaz (O2, CO2), de substances nutritives et de déchets métaboliques, le chorion joue un rôle endocrinien. En effet, il produit deux hormones essentielles au développement embryonnaire:
- hormone chorionique gonadotrophique (HCG): hormone peptidique qui induit la production de progestérone par les cellules maternelles du placenta et, via la circulation sanguine, par l'ovaire maternel. La progestérone conserve la paroi utérine épaisse et vascularisée. Elle est de plus utilisée par les glandes surrénales foetales comme substrat biochimique de la production d'hormones corticostéroïdes. Les tests de grossesse modernes consistent souvent à déceler la présence de HCG dans l'urine maternelle.
- hormone chorionique somatotrophique (lactogène placentaire): hormone qui facilite le développement des glandes mammaires de la mère durant la gestation.
Le chorion protège en plus le foetus contre une réponse immunitaire de la mère. Des antigènes maternels et paternels sont exprimés à la surface membranaire des cellules du trophoblaste (contribution embryonnaire au placenta). Il semble que le chorion produirait des bloqueurs contre une réponse immunitaire maternelle possible.
Qu'est-ce qui, chez les mammifères, détermine la durée de gestation? Bien que réponse à cette question ne puisse être apportée directement, on sait ce qui signale la fin de la gestation. Une élévation de la production d'ACTH (adrenocorticotropic hormone), par l'hypophyse du foetus, induit la production accrue de cortisol par ses corticosurrénales, et cette dernière hormone, par voie sanguine du foetus à la mère, cause un déséquilibre enzymatique dans l'utérus qui résulte en début du travail.
{kind=link}
{kind=link}