> Autres dérivés de l'ectoderme (cours 8)
ORGANOGENESE: NEURULATION ET DERIVES DU NEURECTODERME (cours 7)
Neurulation
La neurulation marque le début de l'organogenèse.
Chez les Vertébrés, la gastrulation a créé un embryon ayant un feuillet ectodermique externe, un feuillet mésodermique intermédiaire et un feuillet endodermique interne. En plus, un cordon de cellules généralement considérées comme mésodermiques, la notochorde, se trouve juste sous la portion dorsomédiane de l'ectoderme. (Certains auteurs considèrent la notochorde, structure transitoire, comme ne faisant partie d'aucun feuillet fondamental; elle constituerait un tissu spécial.) La notochorde exerce un effet inducteur sur l'ectoderme sus-jacent pour qu'il forme le tube neural: il s'agit de l'induction embryonnaire primaire. Nous y reviendrons au dernier cours.
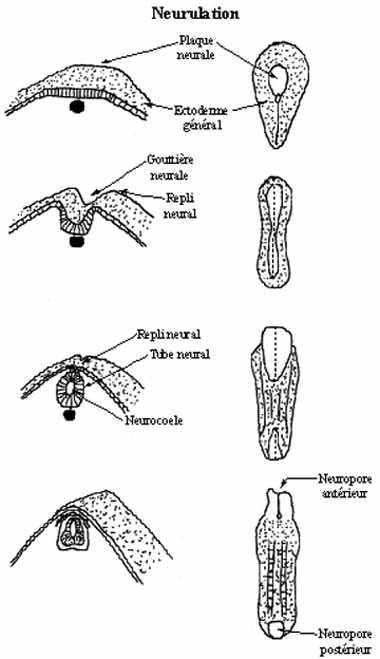
L'embryon sur le point de neuruler s'allonge dans l'axe antéro-postérieur et s'aplatit dorsalement. Le premier indice qu'une région de l'ectoderme devient spécifiée à devenir tissu neural se manifeste par un changement de la forme des cellules de l'ectoderme dorsal de l'embryon. Ces cellules s'allongent, devenant cylindriques, tandis que, de part et d'autre, les autres cellules ectodermiques sont cuboïdales ou pavimenteuses. Les cellules cylindriques dorsales forment ensemble le neurectoderme: épithélium simple (une seule assise de cellules) cylindrique (les cellules qui le composent sont toutes cylindriques). Le neurectoderme étant surélevé par rapport au reste de l'ectoderme (ectoderme général), on l'appelle à ce stade plaque neurale. Près de 50% des cellules ectodermiques sont inclues dans la plaque neurale.
Les rebords de la plaque neurale, à la limite neurectoderme - ectoderme général, s'épaississent et se soulèvent: les replis neuraux, entre lesquels se crée une dépression en forme de U: la gouttière neurale.
En se soulevant les replis neuraux se rapprochent l'un de l'autre, s'attouchent et fusionnent, formant une structure tubulaire, le tube neural, creusée d'une cavité, le neurocoele. Ce faisant, les replis neuraux tirent avec eux l'ectoderme général, qui recouvre maintenant le tube neural. La connexion entre le tube neural et l'ectoderme général se dissout, le tube neural aboutissant entre l'ectoderme général et la notochorde.
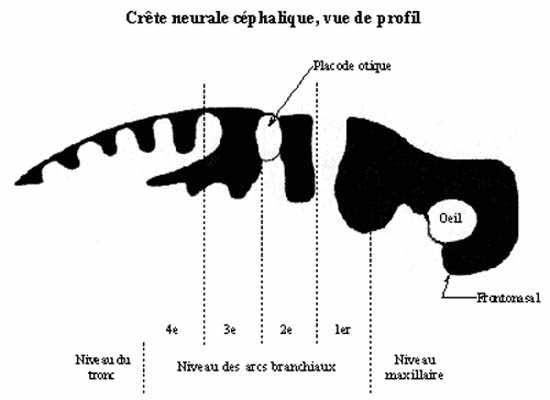
Les cellules comprises dans les replis neuraux ne sont pas incorporées au tube neural, ni ne demeurent soudées à l'ectoderme général. Plutôt, elles se détachent du tube neural en même temps que de l'ectoderme général et forment une structure d'abord unique et médiane, la crête neurale, qui coiffe le tube neural sur toute sa longueur. Sauf au niveau céphalique, la crête neurale se scinde en deux sur la ligne médiane puis se métamérise. Il en résulte des structures segmentées paires.
Le tube neural est à l'origine du névraxe: système nerveux central (cerveau et moelle épinière), et la crête neurale de presque tout le système nerveux périphérique et d'une bonne partie du squelette crânio-facial.
La neurulation ne se produit pas simultanément sur toute la longueur de l'embryon. Amorcée à la jonction tête - tronc, elle se poursuit antérieurement et postérieurement. Chez le poulet de 24 heures d'incubation, elle est fort avancée dans la région de la tête tandis que la région caudale est encore en gastrulation. Chez les amphibiens la neurulation s'effectue en 24 heures à 18°C. Chez les oiseaux elle débute vers la 20e heure d'incubation. L'embryon des Vertébrés ayant subi ces changements s'appelle neurula.
Développement ultérieur du tube neural: SNC
Les deux extrémités du tube neural ne se referment pas immédiatement mais demeurent soudées à l'ectoderme général et ouvertes sur l'extérieur, les neuropores antérieur et postérieur. Chez les amniotes les neuropores permettent le passage de liquide amniotique dans le neurocoele pendant un certain temps. Normalement ils se referment, le postérieur puis l'antérieur. Si le neuropore postérieur ne se referme pas, il en découle une pathologie appelée spina bifida, dont la sévérité dépend de la quantité de moelle épinière demeurant ouverte. Si le neuropore antérieur ne se referme pas, il en résulte une anencéphalie, léthale car le cerveau ainsi exposé dégénère et la voûte crânienne ne se forme pas. L'anencéphalie caractérise 0,1% des grossesses humaines.
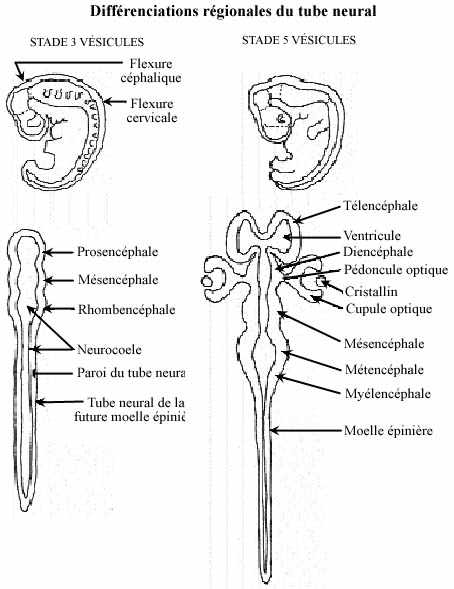
Des changements locaux du tube neural se produisent. Dans la portion céphalique le tube neural, élargi dès sa formation, se resserre en deux endroits et délimite 3 vésicules primaires: prosencéphale, mésencéphale, rhombencéphale. Postérieurement, le tube neural demeure un simple tube s'amincissant caudalement, la future moelle épinière.
Au moment où la portion caudale du tube neural se forme émergent les vésicules optiques, deux évaginations latérales du prosencéphale. Deux pliures ventrales se dessinent: la flexure cervicale, entre le rhombencéphale et la moelle épinière, et la flexure céphalique, entre le mésencéphale et le rhombencéphale. L'embryon devient replié sur lui-même. Ces repliements, du moins cela est connu de la flexure cervicale, ne sont pas intrinsèques au tube neural mais passifs; ils obéissent au mouvement autonome de repliement du coeur, dont nous discuterons dans un autre chapitre.
Par constriction, le rhombencéphale se subdivise en deux: myélencéphale et métencéphale. Le myélencéphale deviendra la moelle allongée, continuation rostrale relativement peu différenciée de la moelle épinière, et le métencéphale donnera naissance au cervelet et structures pontiques associées. Le mésencéphale ne se subdivise pas; il comprendra une portion du tronc cérébral (tegmentum) et le tectum chez les sous-mammaliens ou les corps quadrijumeaux chez les mammifères. Le prosencéphale se subdivise en deux par constriction: diencéphale et télencéphale. Le diencéphale formera le thalamus, le sous-thalamus, l'hypothalamus et l'épithalamus. C'est au diencéphale, plus précisément au thalamus, que sont reliées les vésicules optiques. Le télencéphale formera les noyaux profonds et les hémisphères cérébraux.
Voir les schémas sur la différencation neuronale et le développement de la paroi du tube neural
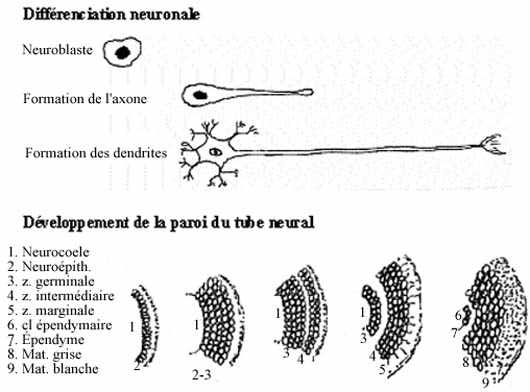
L'épithélium de la plaque neurale, de la gouttière neurale et du tube neural initial est simple cylindrique ou pseudostratifié. Toutes les cellules sont en activité mitotique intense. Les cellules du tube neural se divisent tellement rapidement que l'épithélium devient stratifié. Certaines cellules cessent éventuellement de se diviser et migrent vers la périphérie du tube. La localisation préférentielle des cellules mitotiques et postmitotiques autour du neurocoele définit deux zones dans la paroi du tube neural: zone germinative et zone intermédiaire ou manteau. Les cellules postmitotiques de la zone intermédiaire, encore non différenciées, appartiennent à deux lignées: neuroblastes, précurseurs des cellules nerveuses proprement dites, et glioblastes, précurseurs des cellules gliales. La zone germinative s'amincit à mesure qu'un plus grand nombre de ses cellules deviennent post-mitotiques et enrichissent la zone intermédiaire. La zone germinative se réduit éventuellement à une seule assise de cellules qui deviennent ciliées: l'épendyme. Le neurocoele devient le canal épendymaire dans la moelle épinière et les ventricules dans le cerveau.
Dans la zone intermédiaire, les neuroblastes se différencient en neurones. D'abord, chacun élabore un prolongement cytoplasmique unique, l'axone, dirigé vers la périphérie du tube neural. Le grand nombre d'axones émis à la périphérie du manteau créent une zone additionnelle au tube neural, la zone marginale. Elle s'épaissit à mesure qu'y parviennent les axones des neurones de la zone intermédiaire. Les neurones continuent leur cytodifférenciation, notamment par l'élaboration d'autres prolongements cytoplasmiques, multiples, les dendrites.
Certains glioblastes de la zone intermédiaire migrent vers la zone marginale et se différencient en oligodendrocytes qui forment un manchon isolant autour des axones: la myéline. Ce manchon à forte teneur lipidique donne une apparence blanche à la zone marginale, qui deviendra la matière blanche. Les glioblastes demeurant dans la zone intermédiaire se différencient en astrocytes qui protègent les neurones. La zone intermédiaire devient la matière grise (par "défaut", i.e., par manque de myéline). Les épendymocytes ne se divisent que pour pourvoir au remplacement des cellules gliales. Les neurones mûrs qui meurent ne sont pas remplacés.
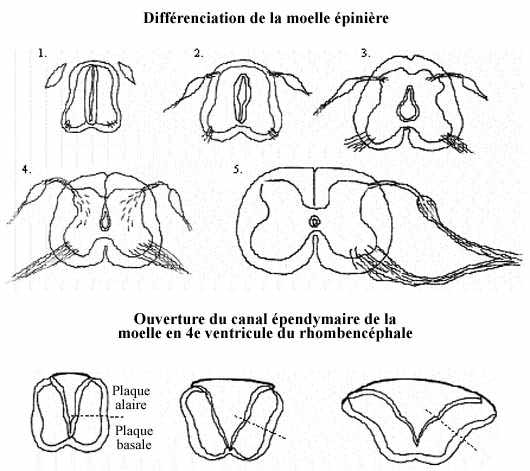
Dans la moelle épinière, le myélencéphale et le métencéphale, les trois zones: épendyme, matière grise et matière blanche, se trouvent concentriques autour du canal épendymaire ou du ventricule. Dans la moelle, la matière grise prend la forme caractéristique d'un H. La moitié dorsale de ces régions, appelée plaque alaire chez l'embryon, est surtout consacrée aux fonctions sensorielles, i.e., la réception de l'information, et devient la corne dorsale. La moitié ventrale, appelée plaque basale, vouée aux fonctions motrices devient la corne ventrale. Les neurones qui s'y retrouvent sont généralement de plus grande taille. La racine ventrale, collection d'axones quittant le SNC pour innerver les muscles squelettiques, prend origine des motoneurones de la corne ventrale. Le terme "corne" est réservé à la moelle épinière.
Le taux des divisions mitotiques puis le mode de migration des neurones post-mitotiques et leur différenciation varient quelque peu d'un niveau à l'autre de la moelle épinière et, pour un niveau donné, tout autour du neurocoele. Ainsi l'épaisseur et la forme des zones intermédiaire et marginale ne sont pas tout à fait les mêmes aux niveau cervical, thoracique, lombaire, sacré ou coccygéen de la moelle, ou bien dans la plaque alaire et la plaque basale.
Dans les régions du cerveau rostrales au métencéphale (més-, di-, tél-encéphale), la façon dont se déroulent les migrations cellulaires et la croissance différencielle du tissu font que les matières grise et blanche s'entremêlent plus ou moins; elles ne sont pas concentriques autour du ventricule. Elles peuvent s'organiser en couches alternées, comme dans le cortex cérébral, le toit optique ou le cortex cérébelleux (ce dernier étant métencéphalique); ailleurs, des neurones à fonction commune se regroupent en amas appelés noyaux, séparés d'autres noyaux à fonctions différentes par des faisceaux d'axones. L'organisation des neurones en noyaux commence déjà à s'observer dans le rhombencéphale.
Les neurones établissent entre eux des contacts fonctionnels spécialisés appelés synapses qui utilisent des médiateurs chimiques appelés neurotransmetteurs.
Les vésicules optiques, évaginations des parois latérales du diencéphale, demeurent rattachées au diencéphale par les pédoncules optiques. Elles sont à l'origine de la rétine de l'oeil. Le développement de l'oeil entier sera étudié au chapitre suivant.
La formation du système nerveux comprend non seulement les étapes de divisions cellulaires, de migration et de différenciation (incluant la formation des synapses et la différenciation chimique - le choix de neurotransmetteurs), mais aussi des étapes de mort cellulaire, i.e., l'élimination sélective d'une proportion des cellules produites, de même que l'élimination d'axones et dendrites ainsi que de synapses établies.
Développement ultérieur de la crête neurale: SNP, squelette crânio-facial
Le développement de la crête neurale signifie grandement celui du système nerveux périphérique. À cause de sa nature périphérique, il nécessite la connaissance du développement et de la constitution adulte des cibles innervées: muscles striés squelettiques permettant les articulations du squelette, muscles lisses et leur organisation en organes, glandes exocrines. Le développement de la crête neurale ne signifie pas que celui du SNP, toutefois. Une grande proportion du squelette de la tête provient en effet de la crête neurale céphalique, non segmentée. Bien que neurectodermique d'origine, nous parlerons de sa formation dans le cadre du développement squeletto-musculaire du mésoderme. Nous nous contenterons, ici, d'énumérer les dérivés de la crête neurale.
Dérivés de la crête neurale:
- cellules nerveuses et gliales des ganglions sensoriels (crâniens et spinaux)
- cellules nerveuses et gliales des ganglions autonomes (sympatiques, parasympathiques)
- médulla de la surrénale
- cellules pigmentées de l'épiderme (mélanocytes, xanthophores, iridophores)
- cellules neuro-endocrines des voies digestives et respiratoires
- cellules parafolliculaires de la thyroïde, sécrétrices de calcitonine
- cellules de Type I du corps carotidien
- odontoblastes de la dent
- tissu conjonctif et squelettique de la tête
- tissu conjonctif des glandes dérivés de l'endoderme et de l'ectoderme pharyngiens
Tout en se divisant, les cellules de la crête neurale migrent vers leur site de résidence définitive. La direction de la migration semble déterminée par le substrat de migration, par contact sélectif des cellules avec la protéine fibronectine. Elles arrêtent leur migration et s'accumulent aux endroits dépourvus de fibronectine.
La destinée des cellules de la crête neurale dépend en partie d'elles mêmes, une détermination intrinsèque, mais aussi de l'environnement: tant du substrat de migration et des autres substances rencontrées lors de la migration que du site de résidence définitive dans l'embryon. Les parts intrinsèque et extrinsèque ne sont pas les mêmes pour toute la crête neurale. Les cellules de la crête neurale sont donc pluripotentes: une cellule peut se différencier en différents types selon sa localisation dans l'embryon; mais toutes n'ont pas le même degré de potentialité dans l'axe antéro-postérieur. Seules celles de la crête neurale céphalique peuvent produire le cartilage de la tête. La crête neurale crânienne transplantée dans la région thoracique formera le cartilage du tronc, qui est normalement produit par le mésoderme. La crête neurale thoracique transplantée dans la région crânienne ne forme pas de cartilage, qui est pourtant normalement formé par la crête neurale en cet endroit. Nous reviendrons sur ces notions au dernier chapitre.
Outre le tissu nerveux même, le système nerveux comprend d'autres tissus. Au cours du développement du tube neural, des cellules mésenchymales locales et le matériel intercellulaire associé forment trois couches autour de lui, les méninges, dont l'épaisseur et la solidité augmentent de l'intérieur vers l'extérieur. La méninge la plus interne, la pie-mère, est la plus délicate et s'insinue dans les fissures du SNC pour lui apporter les capillaires sanguins; la moyenne ou arachnoïde-mère véhicule les vaisseaux sanguins de taille intermédiaire; la dure-mère constitue une capsule conjonctive typique et solide. L'espace sous-dural renferme du fluide et du tissu adipeux qui servent d'amortisseur au SNC. Méninges et vaisseaux sanguins dérivent du mésoderme. Le tissu mésodermique n'entre jamais en contact direct avec les cellules nerveuses, incluant leurs prolongements (axones et dendrites), mais en est toujours séparé par des cellules gliales.
ORGANOGENESE: AUTRES DERIVES DE L'ECTODERME (cours 8)
L'ectoderme nouvellement séparé du tube neural consiste en un épithélium simple pavimenteux à cuboïdal. La plus grande partie de cet ectoderme formera l'épiderme de la peau et les téguments associés, le reste entrera dans la composition des placodes neurogéniques: olfactives, optiques, otiques et lignes latérales, des ganglions sensoriels et autonomes des nerfs crâniens, ainsi que des placodes épibranchiales.
1) Epiderme:
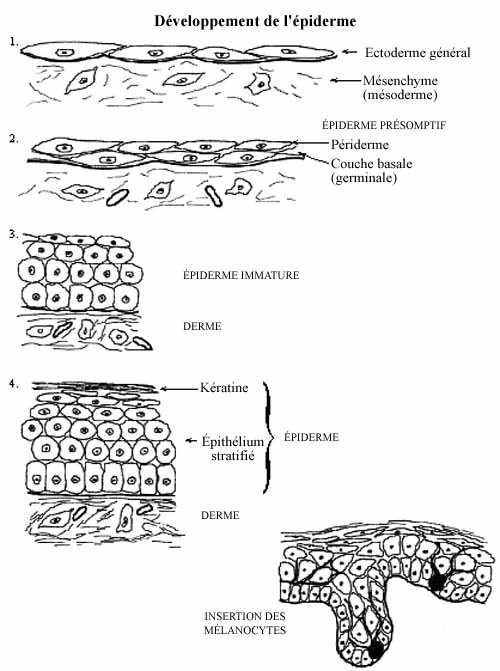
L'épiderme constitue la couche externe de la peau. La peau est le plus gros organe du corps; elle recouvre entièrement l'organisme, le protège du monde extérieur, assure son intégrité et agit comme interface de communication entre le monde externe et l'organisme. L'épiderme de la peau consiste essentiellement en un épithélium stratifié kératinisé. La kératine fait partie intégrante de l'épiderme et consiste en matériel inerte dérivé des cellules épidermiques. C'est la kératine qui assure en grande partie les propriétés de protection attribuées à la peau.
L'épiderme se forme à partir de la simple couche ectodermique qui recouvre l'embryon à la fin de la neurulation. Par divisions mitotiques des cellules de l'épithélium ectodermique, celui-ci devient bistratifié et prend le nom d'épiderme présomptif. Sa couche superficielle, transitoire, s'appelle périderme et sa couche profonde, ou couche basale, est une couche germinative, à partir de laquelle les kératinocytes de l'épiderme seront produits.
Les cellules de la couche germinative entreprennent de nombreuses mitoses. Certaines des cellules-filles demeurent germinatives mais d'autres deviennent post-mitotiques et sont repoussées superficiellement, entre la couche germinative et le périderme. L'épithélium stratifié prend le nom d'épiderme immature. Les cellules postmitotiques commencent à synthétiser les produits nécessaires à la formation de la kératine. L'épiderme ne comprend toutefois pas encore de kératine. Il est toujours recouvert du périderme, dont les cellules ne peuvent mériter le nom de kératinocytes.
À mesure qu'un plus grand nombre de cellules post-mitotiques sont ajoutées à l'épiderme, celui-ci s'épaissit. Comme tout épithélium, même épais, l'épiderme ne renferme aucun vaisseau sanguin; il ne contient que des cellules et ces cellules sont nourries par diffusion à partir du derme sous-jacent (voir plus bas) qui, lui, est vascularisé. Certains kératinocytes deviennent donc de plus en plus éloignés de leur source nutritive dans le derme en formation: ces kératinocytes se transforment, avec les produits de synthèse mentionnés, en kératine. Ainsi, la kératine ne consiste pas uniquement en un produit de sécrétion mais en transformation de cellules entières.
La kératine, un produit inerte, ne demeure pas indéfiniment à la surface de la peau; elle est continuellement desquamée et doit être remplacée. Ainsi, le processus de kératinisation est en branle durant toute la vie de l'animal. La couche basale de l'épiderme demeure couche germinative pendant toute la vie; elle pourvoit au renouvellement des kératinocytes perdus à cause de la kératinisation et la desquamation.
En même temps que se forme l'épiderme, des cellules de la crête neurale migrent vers lui, s'insèrent entre les cellules germinatives de la couche basale (à l'origine des kératinocytes) et se différencient en mélanocytes. Outre la synthèse de mélanine, la différenciation des mélanocytes consiste en l'élaboration de nombreux prolongements cytoplasmiques qui s'insinuent entre les kératinocytes superficiels. Les mélanocytes confèrent à la peau sa pigmentation.
Ce n'est que relativement tard que le périderme embryonnaire est perdu.
Sous l'ectoderme en formation, on retrouve des cellules mésodermiques libres provenant du dermatome des somites qui, avec le matériel intercellulaire qu'elles sécrètent, forment un tissu conjonctif lâche appelé mésenchyme, rudiment du derme. Nous y reviendrons.
2) Téguments et annexes épidermiques:
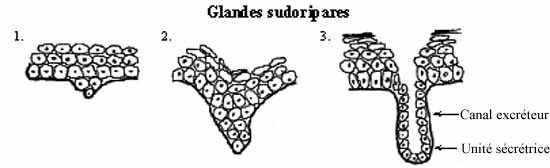
La peau protège l'organisme grâce à son épithélium épais et kératinisé. Ce type d'épithélium ne permet pas l'insertion en son sein de cellules sécrétrices, cellules souvent présentes dans les épithélia et dont la sécrétion assure une protection à l'épithélium qui les abrite: contre la dessication et les microorganismes étrangers. C'est ainsi que, dans le cas de l'épiderme, des glandes profondes sont nécessaires.
Tandis que plusieurs cellules dérivant de la couche germinative sont repoussés vers la surface, certaines cellules, au contraire, prolifèrent profondément dans le mésenchyme sous-jacent. Un canal se creuse à l'intérieur de cette masse cellulaire. La portion distale de ces groupes de cellules profondes se différencie en unité sécrétrice, qui synthétise la sécrétion; l'unité sécrétrice est reliée à la surface de l'épiderme par un canal excréteur qui transporte la secrétion. Unités sécrétrices et canaux excréteurs sont constitués d'un épithélium simple ou bistratifié, cuboïdal à cylindrique, et forment ensemble les glandes sudoripares.
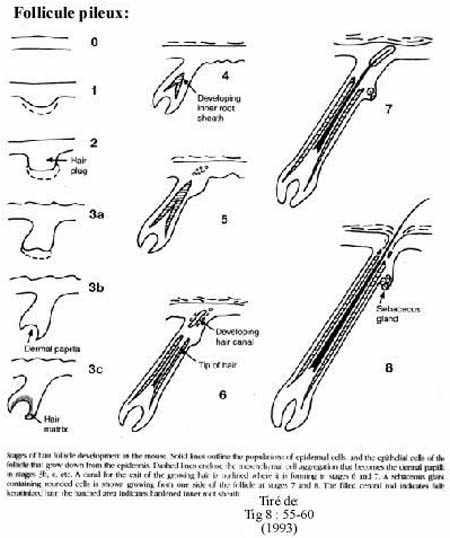
Chez les mammifères, les follicules pileux croissent également par proliférations de l'épiderme dans le derme; cette prolifération se poursuit par une excroissance au travers le derme et l'épiderme, excroissance qui se kératinise, un peu comme la peau mais à un degré plus avancé, et devient le poil.
À partir de la prolifération du follicule pileux se produit une ou plusieurs proliférations secondaires, dans le derme, qui se différencient en glandes sébacées. Ainsi, à chaque follicule pileux est associée au moins une glande sébacée dont la sécrétion, le sébum, lubrifie le poil.
Chez les sous-mammaliens, les téguments diffèrent du poil: plumes, écailles ou autres, mais ils croissent plus ou moins selon le même principe que le follicule pileux.
Chez les mammifères, comme chez les autres, on retrouve d'autres types de téguments: ongles, sabots, cornes, etc. Eux aussi se développent de façon similaire au follicule pileux, par prolifération interne, puis externe, de cellules originaires de l'épithélium germinatif de l'épiderme, puis leur kératinisation.
La kératine, comme on l'a vu, est autant un produit de transformation cellulaire que de sécrétion. Les sécrétions peuvent varier d'un type tégumentaire à l'autre, expliquant la grande diversité des téguments.
Les glandes mammaires, chez les mammifères, se développent sur presque toute la longueur du ventre embryonnaire, formant une structure bilatérale appelée ligne de lait. Elles se forment de façon similaire aux glandes sudoripares mais leurs canaux excréteurs s'embranchent. Leurs unités sécrétrices ne se différencient que chez la femelle adulte durant la gestation. Les primordia des glandes mammaires se retrouvent même chez le mâle mais se résorbent; seule est conservée la portion externe sur la peau. Chez la femelle, une proportion des glandes formées se résorbe, et ne sera conservé que le nombre de glandes mammaires spécifique à l'espèce, à leur localisation spécifique.
3) Placodes neurogéniques:
Placodes des organes sensoriels spéciaux
Certains auteurs considèrent les placodes sensorielles comme étant une contribution ectodermique aux organes récepteurs des systèmes sensoriels spéciaux: olfaction (épithélium olfactif du nez), vision (oeil), audition et équilibre vestibulaire (oreille interne), ainsi que la ligne latérale chez les poissons et les amphibiens larvaires. Nous allons voir qu'elles peuvent être conçues différemment.
Sauf pour la vision, c'est au niveau de la placode que se fait la réception de l'information venant du monde externe: odeur, son, gravité et déplacement.
Le développement de ces différentes placodes se produit de façon similaire. Le rudiment nerveux du sens en question induit dans l'ectoderme immédiatement sus-jacent une prolifération et une différenciation cellulaires qui font que cette portion de l'ectoderme évoluera différemment de la portion adjacente. Cette portion ectodermique élabore un épithélium très spécialisé, fort différent de l'ectoderme de la peau, adapté au sens auquel il est associé.
Comme nous ne reviendrons pas sur ces systèmes sensoriels dans les autres chapitres, nous verrons ici non seulement le développement de la placode mais celui de tout l'organe sensoriel, sans toutefois entrer dans beaucoup de détails.
- Placode cristalline et formation de l'oeil
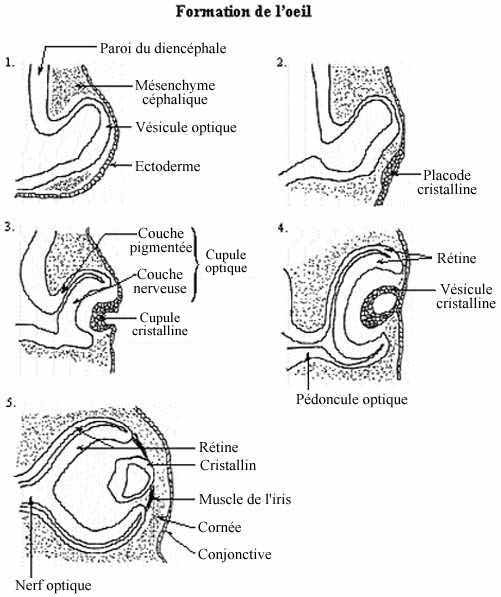
Nous avons vu que les vésicules optiques sont des évaginations des parois latérales du diencéphale, auxquelles elles sont rattachées par les pédoncules optiques. En s'évaginant les vésicules optiques s'approchent de l'ectoderme superficiel, sur lequel elles induisent la formation des placodes optiques ou cristallines. En réalité, l'induction de l'ectoderme appelé à devenir placode cristalline se fait à un stade plus précoce, avant même que la vésicule optique ne soit bien formée, par la portion du tube neural en formation appelée à devenir vésicule optique. Une fois formée, la vésicule optique active cette induction.
La placode cristalline induirait à son tour un changement dans la vésicule optique en la forçant à s'invaginer en cupule optique. La couche "externe" de la cupule optique, ici la plus près du cerveau et la plus éloignée de l'ectoderme, est constituée d'un épithélium simple de cellules pigmentées (mélanine) et forme la couche pigmentée de la rétine, la couche non-nerveuse de la rétine. La portion "interne" de la cupule, un épithélium stratifié, formera la couche nerveuse de la rétine. Dans cet épithélium se retrouvent, entre autres, les photorécepteurs: cônes et bâtonnets. Ainsi, tel que mentionné, dans le cas de la vision, la réception de l'information ne se fait pas par des cellules dérivées de la placode ectodermique mais par des cellules de lignée nerveuse. D'autres cellules de la couche nerveuse de la rétine projettent leurs axones dans le pédoncule optique, formant le nerf optique relié au diencéphale (au thalamus).
En parallèle avec la cupule optique, la placode cristalline s'invagine elle aussi, et dessine la cupule cristalline. Celle-ci se détache de l'ectoderme général et se referme en une structure plus ou moins sphérique: la vésicule cristalline. L'ectoderme général se referme par dessus la vésicule cristalline et la recouvre, comme cela s'était produit après la fermeture du tube neural. La vésicule cristalline se différencie en cristallin de l'oeil. Cette différenciation implique la synthèse de protéines spécifiques, appelées protéines cristallines, synthèse qui obéit à une induction de la rétine nerveuse.
Le cristallin induit l'ectoderme sus-jacent à former la conjonctive, cette mince peau transparente recouvrant l'oeil. Le délicat mésenchyme entre le cristallin et la conjonctive, probablement dérivé de la crête neurale plutôt que du mésoderme, devient la cornée, elle aussi transparente.
Les muscles de l'iris, qui contrôlent la courbure du cristallin et donc l'accomodation de l'oeil, sont uniques en ce qu'ils sont pigmentés, conférant aux yeux leur couleur, et qu'ils dérivent, cela est connu avec certitude, du neurectoderme (plutôt que du mésoderme), plus précisément de replis de la rétine n'ayant pas développé de photorécepteurs.
Les muscles extraoculaires: les deux obliques et les quatre droits, constituent une contribution mésodermique véritable à l'oeil.
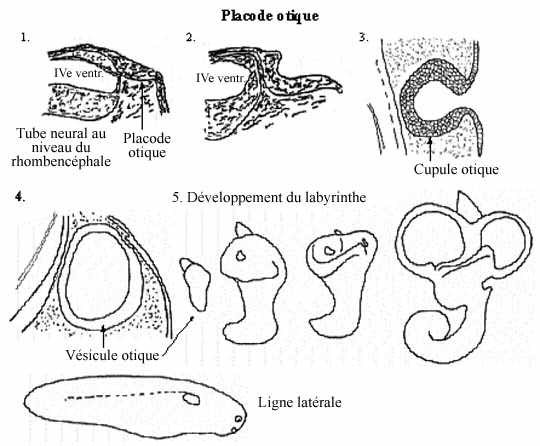
- Placode otique et formation de l'oreille interne et de la ligne latérale
À la jonction myélencéphale - métencéphale, vers le milieu du rhombencéphale, la paroi latérale du tube neural décrit une évagination, bilatéralement. Ces évaginations, moins prononcées que les vésicules optiques s'approchent de l'ectoderme sus-jacent et induisent dans ce dernier la formation des placodes otiques, épaississement ectodermique bilatéral résultant d'une activité mitotique intense.
La placode otique poursuit son développement de façon similaire à la placode optique: elle s'invagine en cupule otique qui se sépare du reste de l'ectoderme et se referme en vésicule otique. Cette dernière se détache de l'ectoderme général qui maintenant la recouvre. La vésicule otique évolue de sorte à former le labyrinthe vestibulaire (utricule, saccule et les trois canaux semi-circulaires): épithélium cilié qui avec des structures accessoires constituent le récepteur de l'équilibre, et la lagéna chez les anamniotes ou la cochlée chez les amniotes: épithélium cilié spécialisé qui avec des structures accessoires constituent le récepteur du son.
Le mésenchyme entourant ces structures épithéliales se condense et se différencie en cartilage ou os formant le squelette de l'organe ainsi qu'en certains muscles associés, de sorte à former l'oreille interne. Nous reparlerons de la nature de ce mésenchyme.
La ligne latérale se développe à partir de cellules de la placode otique qui migrent antérieurement, le long de la tête, de même que postérieurement, sur le flanc de l'embryon. Elles accomplissent une fonction d'équilibre similaire ou complémentaire à celle de l'organe vestibulaire mais ne s'accompagnent pas d'autant de tissu accessoire. La ligne latérale se forme chez tous les embryons anamniotes mais n'est retenue que chez les poissons et certains amphibiens primitifs, pour être résorbée chez les autres. Elle ne se forme pas, ou seulement à l'état très rudimentaire, chez l'embryon des amniotes.
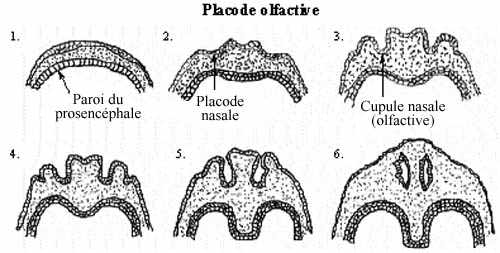
- Placode nasale et organe de l'olfaction
Deux évaginations neuroépithéliales de l'extrémité rostrale du télencéphale induisent l'ectoderme sus-jacent à proliférer et se différencier en placodes olfactives (nasales), puis en cupules, puis vésicules olfactives, épithélium olfactif présomptif composant une partie de la muqueuse nasale, un peu de la même façon que les placodes optiques et otiques se sont formées. Les narines ne s'ouvrent vers l'extérieur que secondairement.
Placodes des ganglions céphaliques
Les placodes neurogéniques ne se retrouvent pas que dans les dérivés des organes des sens spéciaux. Nous avons dit, traitant de la crête neurale au chapitre précédent, qu'elle mène à la formation des ganglions sensoriels et autonomes. La crêtre neurale suffit à la formation des ganglions reliés à la moelle épinière: sensoriels (ganglions spinaux ou rachidiens) comme autonomes (ganglions et plexi sympathiques et parasympathiques). Par contre, elle ne fait que contribuer à la formation des ganglions reliés à l'encéphale.
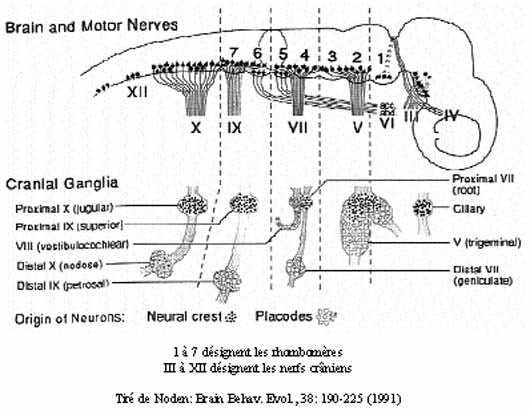
Des placodes, toujours originaires de l'ectoderme de surface, s'associent aux cellules de la crête neurale céphalique pour former les ganglions sensoriels ou autonomes (selon le cas) des nerfs crâniens: ganglion noueux du n. X (vague), ganglion pétreux du n. IX (glossopharyngien); ganglion vestibulo-acoustique du n. VIII (vestibulo-cochléaire), ganglion genouillé du n. VII (facial) et ganglion trijumeau du n. V (trijumeau). Il va sans dire que tous ces ganglions sont des structures paires.
Bien que traitées dans le cadre du chapitre des dérivés ectodermiques, les placodes neurogéniques, comme leur nom le sous-tend, sont réellement des dérivés neuro-ectodermiques. L'on tend de plus en plus à les concevoir comme étant une rétention de tissu neuroépithélial dans l'ectoderme, possiblement spécifié en même temps que la plaque neurale, plutôt que comme une transformation tardive de l'ectoderme général. Ce tissu neuroépithélial finit par se séparer de l'ectoderme général, mais plus tard que ne le fait la plaque neurale.
4) Placodes épibranchiales
Les placodes ectodermiques ne se forment pas qu'en association avec le système nerveux. Des placodes se développent, par exemple, à partir de l'ectoderme recouvrant les arcs branchiaux, ces structures communes à tous les vertébrés embryonnaires et que nous étudierons dans les trois chapitres suivants. Ces placodes se nomment placodes épibranchiales. Il ne semble pas encore être connu si elles surgissent d'elles mêmes ou suite à une induction quelconque. Les placodes des 4ème et 5ème paires d'arcs branchiaux, par exemple, se développeront en glandes parathyroïdes, au nombre de 4 (2 paires). La reconnaissance de l'origine ectodermique de la glande parathyroïde date de relativement peu d'années. On la croyait d'origine endodermique, comme la thyroïde, et c'est l'information que vous retrouvez encore dans plusieurs livres. Nous n'entrerons pas en détail dans ces considérations.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}