ORGANOGENESE: DERIVES DE L'ENDODERME (cours 8)
Les dérivés de l'endoderme sont les suivants:
- épithélium tapissant le tube digestif: bouche et structures associées, oropharynx, oesophage, estomac, petit et gros intestins, de même que les glandes profondes ayant pris origine de l'épithélium tapissant ces organes. Les autres composantes tissulaires de ces organes ne sont pas endodermiques. Bouche et anus comprennent une portion ectodermique et une portion endodermique; la limite entre l'ectoderme et l'endoderme buccal et anal s'avère difficile à tracer.
- épithélium glandulaire des glandes accessoires au tube digestif: foie (et vésicule biliaire, qui n'est pas une glande) et pancréas. Les autres tissus compris dans ces organes ne sont pas endodermiques. En réalité, le pancréas et le foie dérivent d'excroissances de l'épithélium intestinal et la vésicule biliaire d'une excroissance du foie.
- épithélium des branchies et des voies respiratoires: nez, nasopharynx, trachée, bronches, arbre pulmonaire. Les autres tissus composant ces structures ne sont pas endodermiques. Le nez comprend une surface ectodermique (et neurectodermique - la placode olfactive); la limite entre l'ectoderme et l'endoderme nasal s'avère difficile à tracer. La trachée et les portions distales des voies respiratoires sont des excroissances de l'épithélium du pharynx.
- le tissu glandulaire des glandes dérivées de l'épithélium tapissant le pharynx: adénohypophyse, la plus grande partie de la thyroïde et du corps ultimobranchial (ces deux ayant en plus une composante provenant de la crête neurale), une portion du thymus.
Développement du tube digestif même
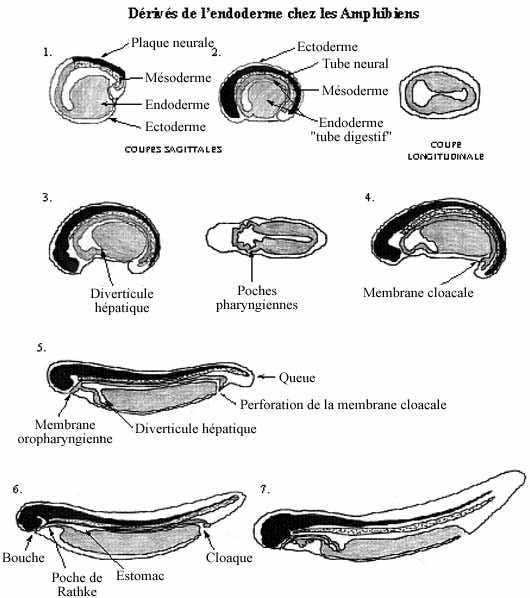
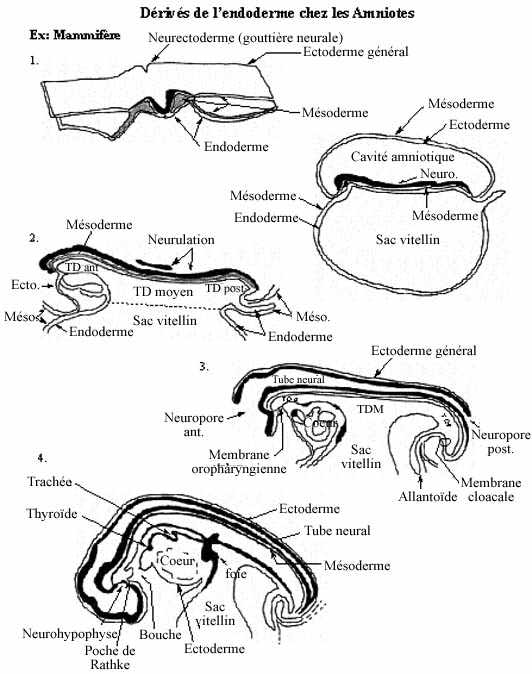
Voir les schémas sur les dérivés de l'endoderme chez les amphibiens et les amniotes
Le tube digestif se forme de façon différente selon que l'embryon dérive d'un oeuf pauvre ou riche en vitellus, donc à segmentation holoblastique ou méroblastique, et selon le type de gastrulation.
Reportons-nous à la fin de la gastrulation. Chez plusieurs poissons et chez les amphibiens, les trois feuillets fondamentaux: endoderme, mésoderme et ectoderme, sont emboîtés, comme des poupées russes; l'endoderme, feuillet le plus interne, renferme une cavité, l'archentéron.
Les trois feuillets concentriques ne sont pas sphériques, à cause de l'élongation antéro-postérieure de l'embryon. Ainsi, l'archentéron tapissé d'endoderme prend une forme tubulaire, présage du futur tube digestif. De plus, son diamètre n'est pas constant sur toute sa longueur. La portion la plus antérieure est relativement élargie et sa paroi endodermique est mince: c'est le tube digestif antérieur. La portion moyenne a un diamètre réduit; la partie dorsale de sa paroi (le toit) est mince tandis que l'épais plancher consiste en blastomères plus fortement vitellins: c'est le tube digestif moyen. La portion caudale est de diamètre réduit et se termine par le blastopore (anus présomptif): c'est le tube digestif postérieur.
En gros, la portion antérieure deviendra l'épithélium tapissant la bouche, le pharynx et la région branchiale, l'oesophage, l'estomac, le duodénum de l'intestin grêle, le foie et le pancréas. La portion moyenne deviendra l'épithélium tapissant le jéjunum et l'iléum de l'intestin grêle et le gros intestin (colons, appendice, rectum). La portion postérieure deviendra le cloaque ainsi que la vessie urinaire, formée à partir du cloaque.
Au site du blastopore, l'endoderme et l'ectoderme ne sont pas séparés l'un de l'autre par du mésoderme. Les deux feuillets fusionnent, formant la membrane cloacale qui se perfore pour devenir l'anus.
À mesure que s'allonge l'embryon et que se caractérisent les différentes régions du tube digestif, certaines d'entre elles s'allongent encore plus que l'embryon entier et deviennent circonvoluées. Ceci est particulièrement marqué dans le cas du tube digestif moyen.
Chez les reptiles et les oiseaux, les 3 feuillets fondamentaux ne sont pas emboîtés à la fin de la gastrulation mais empilés sur le sac vitellin, nonobstant un certain espace entre ce dernier et l'endoderme, espace ne pouvant être considéré comme un archentéron véritable, mais auquel on donne néanmoins le nom, par commodité. Chez les mammifères, l'organisation des feuillets est la même mais le "sac vitellin" ne renferme pas de vitellus. Il a été nommé ainsi par analogie avec le sac vitellin des reptiles et oiseaux.
Le blastodisque se soulève du sac vitellin par un resserrement des feuillets endodermique et mésodermique, comme le resserrement des cordons d'une bourse. Ceci marque la formation de la tige vitelline. Ce faisant, les deux feuillets emprisonnent une portion de l'archentéron, qui mérite maintenant ce nom, car il constitue la lumière du tube digestif en formation. Ce repliement des deux feuillets internes (nous parlerons des mouvements de l'ectoderme plus tard) se produit d'abord dans la région antérieure de l'embryon et définit en premier la portion antérieure du tube digestif. Ensuite c'est la portion postérieure du tube digestif qui s'individualise. La portion moyenne demeure longtemps en communication avec le sac vitellin, car la tige vitelline ne se forme (ne se resserre) que très lentement.
De la région postérieure du tube digestif, caudalement au sac vitellin, émerge ventralement un diverticule endodermique appelé allantoïde, une structure extra-embryonnaire que nous étudierons plus tard. À mesure que le noeud primitif régresse caudalement en fin de gastrulation, il laisse devant lui les trois feuillets, sauf dans la région caudale du blastodisque juste postérieure à l'allantoïde. Il ne se trouve pas de mésoderme entre l'ectoderme et l'endoderme à cet endroit. Donc, endoderme et ectoderme sont directement apposés, puis fusionnent, formant la membrane cloacale. La membrane cloacale s'amincit, se perfore, et crée l'anus, comme chez les anamniotes. Derrière le cloaque s'allonge la queue de l'embryon, recourbée ventralement.
Chez les anamniotes et les amniotes reptiliens et aviaires, le cloaque reçoit les contenus du tube digestif, de l'urètre et des conduits génitaux (mâles ou femelles, selon le sexe). Chez les mammifères, le cloaque se subdivise en deux: un septum croissant à partir de l'allantoïde sépare l'anus du sinus urogénital. Dans le chapitre suivant, l'évolution de cette région chez le mâle et la femelle sera décrite.
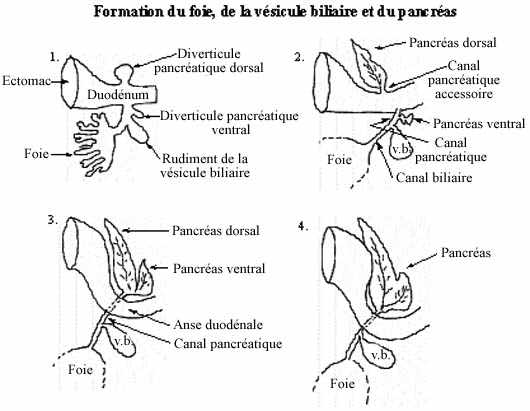
Développement des glandes accessoires au tube digestif (voir schéma)
Une excroissance du plancher du duodénum, le diverticule hépatique, en s'embranchant maintes fois conduit à la formation de l'épithélium glandulaire du foie. À partir de cet épithélium, une autre excroissance produit l'épithélium (non glandulaire) tapissant la vésicule biliaire. La vésicule biliaire sert de réservoir de l'une des sécrétions hépatiques, la bile, et demeure reliée au foie par le canal biliaire. Le foie garde sa communication avec le duodénum, dans lequel il rejette ses sécrétions, par le canal pancréatique.
Une excroissance du toit du duodénum mène à la formation du pancréas dorsal tandis qu'une évagination du plancher du duodénum ou du canal pancréatique donne naissance au pancréas ventral. Sauf chez certains poissons, les deux rudiments pancréatiques se rejoignent éventuellement en un organe unique qui joint le canal pancréatique. Les sécrétions exocrines du pancréas sont donc rejetées dans la lumière duodénale.
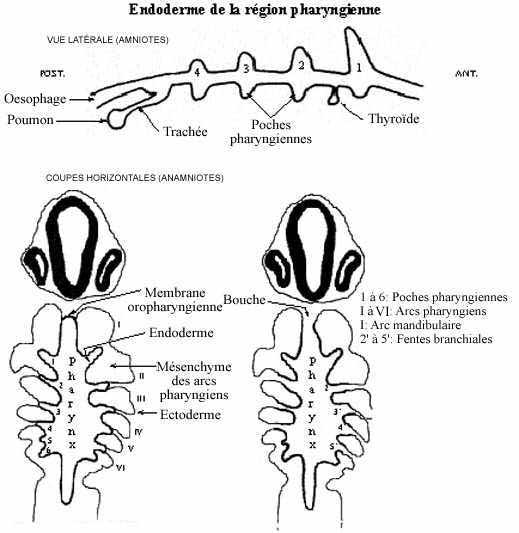
Développement particulier de la région pharyngienne
Au niveau du pharynx, l'archentéron est élargi mais il est également aplati. Au point rostral du pharynx, ventralement, là aussi l'endoderme entre en contact avec l'ectoderme général et ce dernier s'affaisse en cet endroit. Les deux épithélia fusionnent et forment la membrane oropharyngienne (ou stomodéale). Comme dans le cas de la membrane cloacale, la membrane oropharyngienne s'amincit, puis se perfore, résultant en la formation de la bouche ou stomodeum. Le tube digestif est maintenant en communication avec le monde externe par les deux extrémités.
Latéralement, la paroi du pharynx décrit des évaginations digitées, les poches pharyngiennes (ou branchiales). Elles sont au nombre de 6 paires chez les anamniotes et de 4 paires chez les amniotes, et se développent selon un gradient antéro-postérieur. Elles se trouvent à délimiter les arcs pharyngiens (ou branchiaux ou viscéraux).
En s'évaginant, la poche pharyngienne repousse le tissu qui la sépare de l'ectoderme; ce dernier s'invagine et entre en contact avec l'endoderme de la poche pharyngienne; les deux fusionnent et forment la membrane branchiale. Sauf pour la 1ère et la 6ème paires, les membranes branchiales des anamniotes se perforent, résultant en la formation des fentes branchiales. Chez les Vertébrés aquatiques, des filaments branchiaux se développent à la surface des fentes branchiales; ce sont les branchies, surface respiratoire de ces animaux. Chez les amniotes, les poches peuvent ne pas du tout s'ouvrir vers l'extérieur; celles qui se perforent se referment secondairement. Comme des branchies ne se développent pas chez les amniotes, certains auteurs préfèrent éviter le terme branchial pour qualifier tant les poches que les arcs, et utiliser le terme pharyngien qui peut s'appliquer à tous les Vertébrés.
Chez les amniotes, la première paire de poches pharyngiennes évolue en trompes d'Eustache. La seconde paire sera incorporée à l'amygdale pharyngienne. Le tissu de la troisième paire se détache du reste du pharynx, migre postérieurement et médialement et se différenciera en thymus. Celui de la quatrième paire devient les corps ultimo-branchiaux, auquel s'ajoutent des cellules migrant de la crête neurale céphalique.
La glande thyroïde (ses cellules folliculaires) se développe à partir de la paroi ventrale et médiane du pharynx, plus précisément de la région juste caudale à la 1ère paire de poches. Les cellules parafolliculaires et le tissu conjonctif de la thyroïde, comme celui des autres glandes dérivées du pharynx, proviennent de la crête neurale céphalique.
Au niveau du pharynx correspondant au diencéphale sus-jacent, l'épithélium dorsal du pharynx dessine une petite évagination, la poche de Rathke, rudiment de la portion glandulaire de l'hypophyse ou adénohypophyse. En s'évaginant, la poche de Rathke rencontre la paroi ventrale de l'hypothalamus qui s'évaginait: l'infundibulum ou éminence médiane; c'est le rudiment de la portion neurale de l'hypophyse ou neurohypophyse. Les éléments endodermique et neurectodermique s'associent intimement, sans cependant fusionner, et forment ensemble l'organe appelé hypophyse (ou pituitaire).
Dorsalement et latéralement au stomodéum, au niveau où se forment les placodes olfactives, une invagination ectodermique en forme de U entre en contact avec l'endoderme pharyngien et indique la formation du nez, dont l'ouverture paire deviendra les narines. L'organe olfactif est incorporé aux narines.
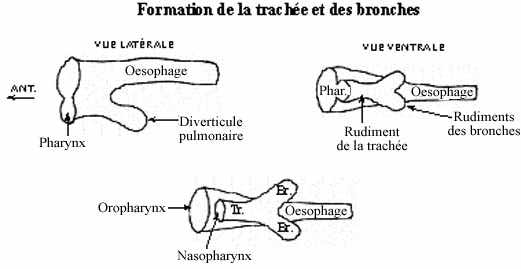
Développement des voies respiratoires (voir schéma)
Postérieurement au pharynx, une évagination du plancher du tube digestif antérieur se dessine. Ce rudiment de la trachée bifurque en rudiments des bronches. Les rudiments des bronches s'allongent et se ramifient, leurs ramifications elle-mêmes s'embranchant et ainsi de suite. Ces embranchements des bronches et le tissu mésodermique qui leur est associé (dont des vaisseaux sanguins) constituent les poumons. La vessie natatoire des poissons se développe de façon comparable, mais sans ramifications.
Le pharynx se divise partiellement en deux portions par un septum: le nasopharynx communique avec la cavité nasale d'une part et la trachée d'autre part, l'oropharynx communique avec la cavité orale d'une part et l'oesophage d'autre part.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}