> Dérivés du mésoderme (cours 9)
> Système cardiovasculaire (cours 10)
ORGANOGENESE: DERIVES DU MESODERME 1 (cours 9)
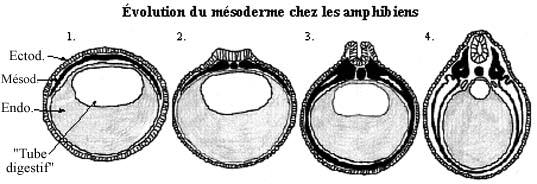
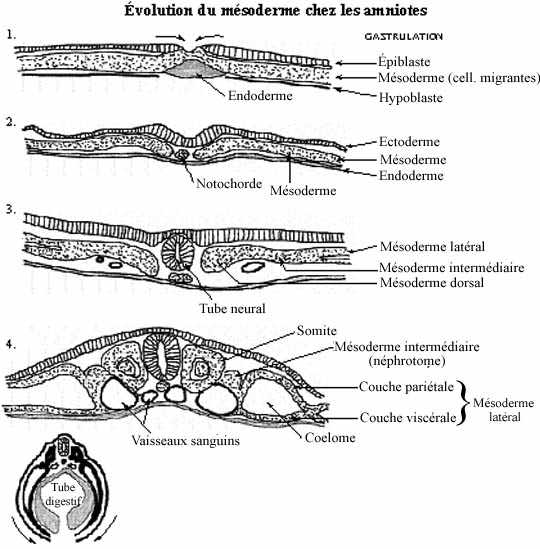
Voir les schémas sur l'évolution du mésoderme chez le amphibiens et les amniotes
À l'issue de la gastrulation, chez les anamniotes, le mésoderme est emboîté entre l'endoderme tubulaire et l'ectoderme, et forme la plaque chordamésodermique, plus épaisse dans la région dorsale que dans les régions latérale ou ventrale de l'embryon. D'ailleurs, la migration des cellules mésodermiques jusqu'au point ventral de l'embryon n'est pas achevée. Chez les amniotes, les feuillets fondamentaux sont empilés et l'archentéron n'est pas encore délimité mais le deviendra fort graduellement par la formation de la tige vitelline.
Très tôt, la région dorsomédiane de la plaque chordamésodermique se condense et se détache du reste de la plaque: c'est la notochorde, présente du niveau du mésencéphale jusqu'au niveau du cloaque, environ, sous le tube neural en formation.
Au niveau du tronc, le mésoderme situé de part et d'autre de la notochorde se distingue en trois portions: mésoderme dorsal, mésoderme intermédiaire et mésoderme latéral. Le mésoderme dorsal, séparé de la notochorde mais encore rattaché au mésoderme intermédiaire, commence à se segmenter, indice de la formation des somites. En se métamérisant, les somites se séparent graduellement du reste du mésoderme mais demeurent rattachés entre eux dorsalement pendant un certain temps. Le mésoderme intermédiaire lui aussi tend à se segmenter, mais un peu plus lentement que les somites et de façon moins marquée. Le mésoderme latéral demeure non segmenté et rattaché temporairement au mésoderme intermédiaire.
La différenciation du mésoderme en ces trois portions débute dès que s'amorce la neurulation à un segment donné et obéit à un gradient médio-latéral. Nous verrons que le mésoderme de la tête se différencie autrement. Donc, la segmentation du mésoderme se poursuit selon un gradient rostro-caudal à partir de la région pharyngienne. Pendant ce temps l'embryon s'allonge antéro-postérieurement et s'aplatit dorso-ventralement sans toutefois augmenter considérablement son volume total. Sa portion antérieure est plus large que la postérieure. La région postérieure au cloaque s'allonge et définit le rudiment de la queue, dans lequel les trois feuillets fondamentaux ne sont pas encore distincts histologiquement. L'âge embryonnaire est souvent exprimé en nombre de somites.
Au niveau céphalique, le mésoderme diffère. Rostralement aux somites et jusqu'au niveau de la vésicule optique, se forme par condensation le mésoderme parachordal qui, comme l'indique son nom, se situe de part et d'autre de la notochorde et un peu plus rostralement. Il consiste en "somites" incomplètement segmentés, auxquels on donne le nom de somitomères, pour éviter la confusion avec les somites véritables. On compte 7 paires de somitomères, numérotées de rostral à caudal. Nous aurions pu mentionner, dans le cadre du développement du système nerveux, que le rhombencéphale aussi se métamérise incomplètement en 7 ou 8 paires de rhombomères, transitoires. Contrairement à la moelle épinière, dont la "métamérisation" (incomplète) succède à celle des somites, la métamérisation (incomplète) du rhombencéphale précède celle des somitomères.
Franchement rostral à la notochorde, le mésoderme préchordal chevauche partiellement le mésoderme parachordal, médian aux deux premiers somitomères, et s'étend rostralement sans toutefois atteindre le point le plus rostral de la tête.
Dans la tête, le mésoderme intermédiaire est inexistant au niveau du mésoderme parachordal et préchordal. On retrouve du mésoderme latéral, scindé en deux feuillets par un coelome, comme dans le tronc, mais ne se rendant pas très rostralement.
Le reste des cellules occupant l'espace entre les feuillets ectodermique et endodermique, au niveau de la tête et particulièrement dans sa région la plus rostrale, et qui subissent un sort comparable aux cellules mésodermiques, dérivent de la crête neurale. Ce tissu interstitiel porte le nom de mésenchyme.
Le terme mésenchyme désigne le tissu interstitiel, retrouvé un peu partout dans l'embryon, remplissant les espaces entre les feuillets et structures bien définis. Peu abondant dans le très jeune embryon, il augmente au cours du développement. Le mésenchyme possède une certaine pluripotentialité, pouvant se différencier en tous les types cellulaires dérivés du mésoderme: cellules conjonctives, chondrogéniques, ostéogéniques, sanguines et musculaires. La plupart se différenciera en tissu conjonctif appelé "ordinaire" chez l'individu mûr. On l'appelle souvent tissu conjonctif embryonnaire. Dans l'embryon, le mésenchyme consiste surtout en cellules mésodermiques qui ne se sont pas condensées. Dans la région de la tête, le mésenchyme dérive en plus de cellules de la crête neurale qui, au lieu de se différencier sur une lignée neurogénique, se différencient comme les cellules mésodermiques.
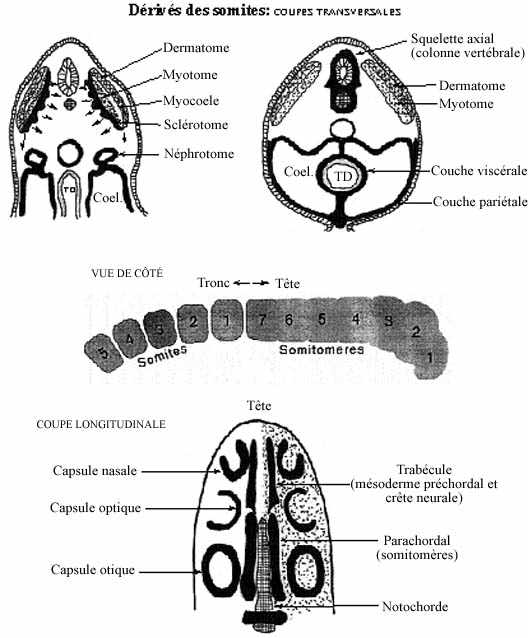
1) Dérivés des somites et somitomères (voir schéma)
Le somite initial consiste en une masse cellulaire peu dense, creusée d'une étroite cavité, le myocoele. Chaque somite s'allonge dorso-ventralement et s'aplatit médio-latéralement. La région dorso-latérale du somite est le dermatome, à l'origine du derme de la peau. La région du somite ventro-médiane au myocoele comprend deux composantes: le myotome, à l'origine des muscles squelettiques, et le sclérotome, dont dérive le squelette et certains muscles squelettiques. On retrouve l'équivalent des dermatome, myotome et sclérotome dans les somitomères.
- Dermatomes: derme de la peau
Les cellules des dermatomes se détachent des somites et somitomères et migrent, tout en se divisant mitotiquement pour augmenter leur nombre, jusque sous l'ectoderme général. Celui-ci forme l'épiderme. Elles se différencieront en cellules du tissu conjonctif qui produisent les fibres conjonctives ainsi qu'en cellules musculaires lisses, le tout formant la couche profonde de la peau: le derme. Ainsi, la peau a une origine embryonnaire double: l'épiderme dérive de l'ectoderme et le derme du mésoderme.
Ce n'est pas le derme de toute la peau du corps, toutefois, qui provient des dermatomes. Le derme d'une bonne partie de la tête provient du mésenchyme de la tête dérivant de la crête neurale céphalique, non métamérisée.
- Sclérotomes: squelette axial et paraxial et partie du squelette de la tête
Les cellules des sclérotomes des côtés gauche et droit migrent autour du tube neural et de la notochorde, les enveloppent et forment le tube périchordal, initialement continu. Selon un gradient rostro-caudal, le tube périchordal se différencie en régions de forte densité cellulaire alternant avec des régions de faible densité cellulaire. Les régions denses deviendront les disques intervertébraux et celles moins denses les corps des vertèbres, auxquels se fixent les côtes. La notochorde disparaît graduellement, quoi qu'il semble maintenant établi qu'une portion demeure et s'incorpore aux corps vertébraux ainsi qu'aux disques intervertébraux. Les sclérotomes conduisent donc à la formation du squelette axial (vertèbres) et paraxial (côtes), mais pas entièrement à celui de la tête.
Le squelette de la tête peut être divisé en quelques régions dérivant de précurseurs variés qui s'agencent pour former la boîte crânienne. Les sclérotomes des 5 premiers somites ainsi que ceux des somitomères (mésoderme parachordal), d'origine mésodermique véritable, participent à la formation des os crâniens, particulièrement ceux de la base du crâne. Plus rostralement, au niveau du diencéphale et du télencéphale, les trabécules proviennent du mésoderme préchordal pour leur portion postérieure et, surtout, de la crête neurale pour le reste. Latéralement à ces deux régions, les capsules des organes sensoriels dérivent en partie du sclérotome de la tête et, surtout, de la crête neurale. En croissant, les trabécules fusionnent ventralement au prosencéphale, puis avec les sclérotomes parachordaux; les capsules des organes des sens rejoignent ces derniers et fusionnent avec eux. La croissance totale de tous ces éléments fait qu'ils viennent à entourer entièrement l'encéphale. Ainsi, le squelette de la tête a une origine double: mésodermique et neurectodermique. Les os formant le maxillaire et la proéminence fronto-nasale dérivent entièrement de la crête neurale.
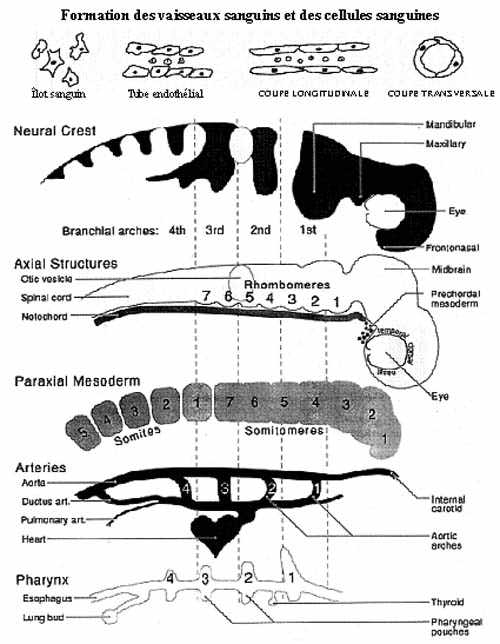
Nous avons vu au chapitre précédent la formation des poches pharyngiennes et leur développement possible en fentes branchiales. Les poches et les fentes délimitent des structures solides: les arcs pharyngiens (ou branchiaux ou viscéraux - deux qualificatifs moins exacts). Leur nombre correspond à celui des poches: 6 paires chez les anamniotes et 4 paires chez les amniotes. Ils consistent en mésenchyme délimité sur la face externe par l'ectoderme et sur la face interne par l'endoderme pharyngien. Le mésenchyme des arcs pharyngiens consiste en mésoderme de la tête auquel s'ajoutent des cellules migrantes de la crête neurale céphalique.
La 1ère paire d'arcs pharyngiens, les arcs mandibulaires, se situe antérieurement à la 1ère paire de poches et entoure le stomodeum, qui devient la bouche. La portion osseuse de cette paire d'arcs formera la mandibule en plus, chez les amniotes, d'osselets de l'oreille moyenne. Différant peu du patron embryonnaire chez les anamniotes, les autres arcs pharyngiens voient leur développement considérablement modifié chez les amniotes. La composante squelettique de la 2e paire d'arcs, les arcs hyoïdiens, formera l'étrier et le styloïde, et celle des deux paires caudales les os et cartilages hyoïdien et thyroïdien.
- Myotomes: Muscles striés squelettiques
Originellement, chaque myotome, l'un à la suite de l'autre, devient un muscle strié squelettique, ou une portion de tel muscle, dans l'axe antéro-postérieur de l'embryon. D'une localisation d'abord dorsale, ils s'étendent ventralement entre le dermatome de la peau en formation et le mésoderme latéral. La portion dorsale du myotome: l'épimère, s'attache aux côtes dorsales et vertèbres du squelette; la portion ventrale: l'hypomère, s'attache aux côtes ventrales et sternum. Elle rejoint l'hypomère de l'autre côté et les deux fusionnent sur la ligne médiane. La musculature du tronc est ainsi formée et c'est le développement maximal qu'atteignent les myotomes chez Amphioxus, les poissons et les larves d'amphibiens.
Chez les Vertébrés terrestres, tétrapodes, l'apparition des membres et le changement du mode de locomotion coincident avec une modification de la segmentation des muscles squelettiques.
Chez les amphibiens, la métamérisation du mésoderme dorsal (épimère et hypomère des myotomes) s'effectue presque jusqu'au niveau du rostrum, car le cou et la tête ne sont pas considérablement développés. Chez les amniotes, le développement important du cerveau résulte en la formation d'une tête considérable. Le développement des muscles striés squelettiques de la tête suit un cheminement parallèle au développement des éléments du squelette auxquels ils sont rattachés. Des cellules des myotomes des 5 premiers somites et des somitomères (mésoderme parachordal) formeront les muscles du cou et de la face. Les muscles de la langue proviennent de cellules des myotomes des 5 premiers somites ayant migré rostralement et sous le pharynx. Les muscles oculomoteurs (les 2 obliques et les 4 droits) proviennent de cellules des myotomes de somitomères (1,2,3,5) ayant elles aussi migré rostralement.
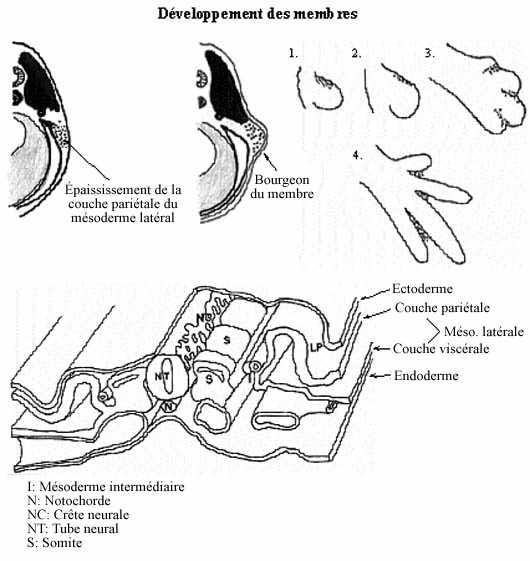
- Développement squelettique et musculaire des membres (voir schéma):
Le squelette et la musculature (striée squelettique) des membres proviennent des sclérotomes et myotomes de certains somites, mais entre aussi dans la composition des membres une portion du feuillet pariétal du mésoderme latéral et, bien entendu, le membre est recouvert de peau (composantes mésodermique et ectodermique).
Le premier indice de la formation d'un membre est un épaississement de la partie supérieure du feuillet pariétal du mésoderme latéral. Les cellules de cet épaississement se détachent du feuillet et forment une masse mésenchymale qui migre latéralement sous la peau en formation. Chez les amphibiens, ces épaississements se retrouvent en positions des futurs membres: aux niveaux brachial et pelvien, bilatéralement. Chez les autres tétrapodes, ils peuvent se produire sur presque toute la longueur du tronc mais ne continuent de se développer qu'aux endroits des membres, régressant ailleurs. Les cellules des sclérotomes et des myotomes migrent vers les épaississements qui font maintenant saillie à la surface: ce sont les bourgeons des membres. Les cellules du bourgeon se divisent plus rapidement que celles adjacentes du tronc de l'embryon. À l'intérieur du bourgeon, les cellules se divisent plus rapidement dans la portion distale que proximale, le bourgeon croissant ainsi plus rapidement par son extrémité.
Quand la longueur du bourgeon excède sa largeur, la différenciation interne du membre commence. Le mésenchyme central, originaire des sclérotomes, se différencie en squelette, d'abord cartilagineux puis osseux, selon un gradient proximo-distal. Le mésenchyme qui l'entoure, provenant des myotomes, s'allonge et se différencie en muscles striés, selon le même gradient. Les cellules du mésoderme latéral fournissent probablement la composante conjonctive. Les vaisseaux sanguins se forment in situ (voir chapitre suivant). L'innervation motrice des muscles provient des neurones moteurs somatiques de la corne ventrale (plaque basale) de la moelle épinière; elle est très précoce.
Le bourgeon du membre s'aplatit, surtout distalement. La région distale aplatie s'élargit et, de circulaire, devient pentagonale, les pointes représentant les bourgeons des doigts. Les bourgeons des doigts continuent de croître et le tissu entre eux se nécrose, menant à la formation de doigts bien individualisés, sauf chez les animaux à pattes palmées chez qui la nécrose est moins accentuée.
Une flexure se crée à l'origine du membre, l'articulant avec le tronc: l'épaule pour le membre antérieur, la hanche pour le membre postérieur. Vers le milieu du membre, une autre flexure se dessine et détermine la position du joint du coude ou du genou qui sépare le bras ou la cuisse de l'avant-bras ou la jambe. Une flexure plus distale marque le joint du poignet ou de la cheville qui sépare l'avant-bras de la main ou la jambe du pied. De plus, les mains et les doigts de même que les pieds et les orteils s'articulent en plusieurs segments par des jointures additionnelles.
Originellement, les muscles fléchisseurs des membres se retrouvent ventralement et les muscles extenseurs dorsalement. Au fur et à mesure de la croissance, il se produit, chez plusieurs tétrapodes, une rotation de l'épaule et de la hanche qui résulte en des positionnements postérieur des fléchisseurs et antérieur des extenseurs.
Notons que les nageoires pectorales des poissons sont les précurseurs évolutifs des membres des tétrapodes et que leur formation implique un apport du feuillet somatique du mésoderme latéral. Nous n'entrerons pas dans ces détails.
2) dérivés du mésoderme latéral: mésentères et muscles lisses
Très tôt, et avant même que la migration ventrale soit terminée, le mésoderme latéral se creuse d'une cavité appelée coelome général, qui le divise en feuillets pariétal (somatique) et viscéral (splanchnique). La cavité coelomique est initialement continue sur toute la longueur de l'embryon. Au cours du développement, des replis du feuillet somatique appelés mésentères la partitionnent en cavités multiples:
- la plèvre tapisse la cavité pleurale et enveloppe tout le thorax
- le péricarde tapisse la cavité péricardiale et enveloppe le coeur
- le péritoine tapisse la cavité péritonéale et enveloppe l'abdomen
La cavité abdominale est séparée de la cavité thoracique par un septum additionnel, le septum transverse, dans lequel migrent des cellules des myotomes qui formeront le muscle strié squelettique qu'est le diaphragme.
La couche mésodermique viscérale se différencie en muscle lisse et tissu conjonctif qui s'associent aux organes internes (organes des systèmes digestif, respiratoire, uro-génital) et aident à leur fonction respective (broyage des aliments pour l'estomac, mouvement péristaltique pour l'intestin, l'uretère et l'oviducte, etc.).
Nous verrons au chapitre suivant qu'une région particulière du mésoderme latéral, ventrale au pharynx, forme le coeur.
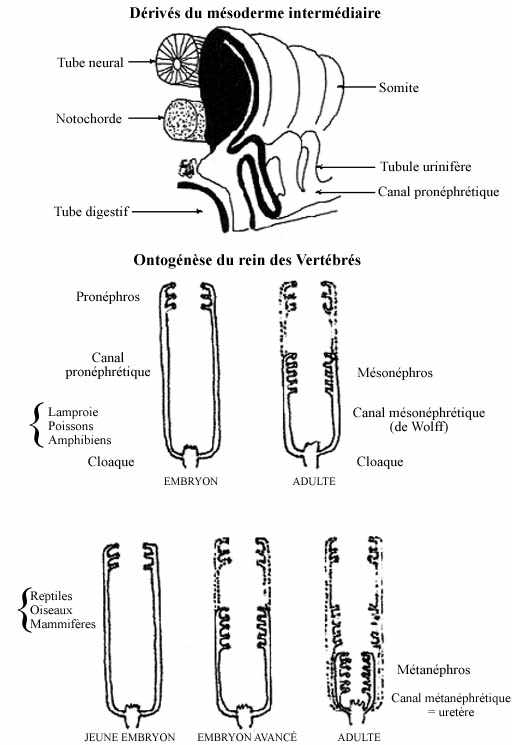
3) Dérivés du mésoderme intermédiaire: système uro-génital (voir schémas)
- Système urinaire:
Initialement, le mésoderme intermédiaire est continu antéropostérieurement, et est ainsi appelé cordon néphrogénétique. Comme on l'a dit auparavant, il se forme selon un gradient antéropostérieur et est d'abord rattaché aux somites, desquels il se sépare tôt, et au mésoderme latéral, duquel il se détache plus tard. Le cordon néphrogénique est plus court que la notochorde ou les somites, ne débutant qu'au niveau caudal du pharynx, et sa formation progresse lentement caudalement. Il se délamine en deux couches: pariétale et viscérale, séparées par le néphrocoele, continu avec le myocoele du somite, transitoirement, et avec le coelome général du mésoderme latéral.
Le cordon néphrogénique se métamérise; chaque segment s'appelle néphrotome. Chaque néphrotome s'allonge considérablement par divisions mitotiques actives des cellules de sa paroi pariétale: on l'appelle tubule urinifère ou tubule rénal. La portion du tubule constituée du feuillet viscéral entre en contact intime avec un réseau de capillaires sanguins appelé glomérule (formé depuis le mésenchyme local). La couche viscérale du tubule et le glomérule forment ensemble le corpuscule rénal. C'est au contact de l'épithélium capillaire (endothélium) et de l'épithélium viscéral du tubule rénal (donc le corpuscule) que s'accomplit la fonction de filtration du plasma sanguin, i.e., la fonction urinaire.
La connexion entre le néphrocoele et le coelome général demeure chez les Vertébrés inférieurs. Ainsi, chez les cyclostomes et certains poissons, dans chaque métamère il se développe un tubule urinifère à partir d'un néphrotome et chacun excrète son produit urinaire dans le coelome. Chez les animaux supérieurs, cette connexion est perdue. Les néphrocoeles des tubules urinifères établissent alors une communication entre eux, à leur extrémité distale, créant un canal excréteur commun: le canal pronéphrétique.
L'organe entier constitue le pronéphros et implique dans sa formation environ le tiers antérieur du cordon néphrogénétique. Les tubules rénaux (pronéphrétiques) et le canal pronéphrétique s'allongent considérablement. Les tubules rénaux, en conséquence, deviennent extrêmement contournés. Le canal pronéphrétique, droit, croît postérieurement dans l'embryon jusqu'au cloaque, dans lequel il se jette. C'est ainsi que les déchets urinaires peuvent être acheminés à l'extérieur de l'organisme.
Bien que fonctionnel chez l'embryon anamniote, le pronéphros se développe chez l'amniote sans jamais être fonctionnel. Les tubules pronéphrétiques dégénèrent mais la portion du canal pronéphrétique caudale au pronéphros demeure. Pendant que dégénère le pronéphros, le mésonéphros se forme à partir du tiers moyen (environ) du cordon néphrogénique, de façon similaire au pronéphros et en association avec des capillaires sanguins (glomérules). Les néphrocoeles de ces tubules rénaux (mésonéphrétiques) entrent en communication avec des ramifications du canal pronéphrétique; ce dernier devient canal mésonéphrétique (ou de Wolff).
La partie antérieure du mésonéphros dégénère chez plusieurs, le reste demeure et constitue le rein fonctionnel des poissons et amphibiens adultes. Chez les reptiles et les oiseaux, le mésonéphros fonctionne chez l'embryon, jusqu'à l'éclosion. Chez l'embryon mammifère, le placenta se charge de l'élimination des déchets métaboliques.
Le rein définitif des amniotes est le métanéphros, qui se développe de façon similaire, à partir de la portion postérieure du cordon néphrogénique, adjacente au cloaque. Le métanéphros conserve le segment postérieur du canal mésonéphrétique en guise de canal excréteur, très court et tout près de sa jonction avec le cloaque; on l'appelle maintenant canal métanéphrétique ou uretère. Le segment antérieur du canal mésonéphrétique dégénère chez la femelle, mais chez le mâle il sera incorporé au système reproducteur, comme nous verrons bientôt.
Le canal métanéphrétique (l'uretère) s'allonge et se jette dans un renflement du cloaque appelé vessie urinaire. À proximité du métanéphros, l'uretère s'embranche, et chaque branche forme un tubule collecteur qui se rattache à plusieurs tubules rénaux. Nous parlerons de la subdivision du cloaque plus loin.
Le rein retient sa position caudale chez les reptiles mais migre antérieurement chez les oiseaux et les mammifères, moyennant un allongement de l'uretère.
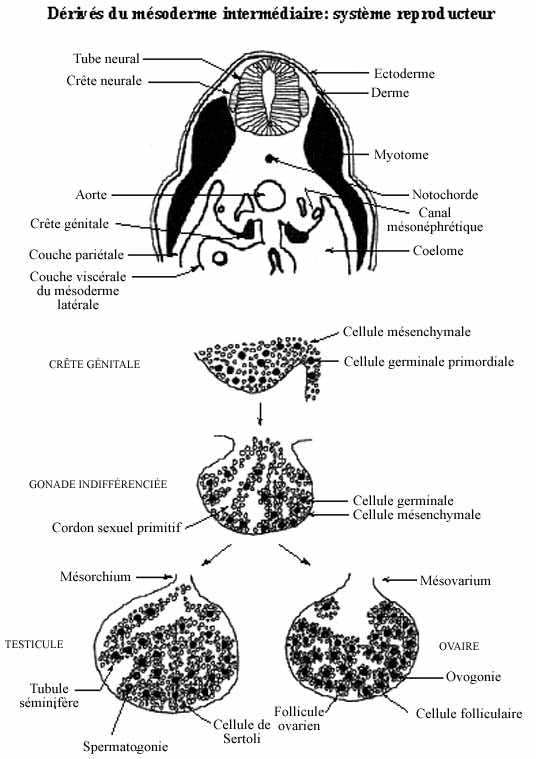
- Système reproducteur (voir schéma):
Comme nous avons vu dans les chapitres traitant de la gamétogenèse, le système reproducteur comprend deux composantes principales: les gonades et une série de canaux excréteurs auxquels s'associent des glandes.
Les gonades proviennent d'un épaississement de la région supérieure du mésoderme viscéral, au niveau du mésonéphros: la crête génitale (germinale). La crête génitale fait saillie dans le coelome, rattachée au mésoderme viscéral par un court mésentère. Initialement composée de cellules du mésoderme viscéral, la crête génitale devient colonisée par un second type cellulaire: les cellules germinales primordiales (gonies), qui s'insèrent parmi les cellules mésodermiques.
Les cellules germinales ont migré à partir de l'endoderme du tube digestif (anamniotes) ou du sac vitellin (amniotes) mais n'en dérivent pas. Elles se sont apparemment différenciées très tôt, durant l'étape de clivage, et se sont insérées dans l'hypoblaste puis l'endoderme. Les cellules germinales ne sont donc pas considérées comme appartenant à l'un ou l'autre des trois feuillets primordiaux de l'embryon, ceux-ci ne générant que les cellules somatiques; elles constituent une lignée propre, la lignée germinale.
Le mésoderme latéral initial contribue peu au tissu de la future gonade. Des cellules du cordon mésonéphrétique qui ne participent pas à la formation du rein migrent jusque dans la crête génitale, où elles entourent les cellules germinales primordiales. Elles se différencieront en cellules de soutien, probablement sous l'effet inducteur des cellules germinales. Les deux types cellulaires forment ensemble les cordons sexuels primitifs. La crête génitale ainsi constituée prend le nom de gonade indifférenciée.
Chez le mâle, les cordons sexuels se creusent d'une cavité, devenant tubulaires: les tubules séminifères. Leur paroi est constituée de cellules provenant du cordon mésonéphrétique: les cellules de Sertoli, et des cellules germinales primordiales: les spermatogonies. Au sortir du testicule, les tubules séminifères deviennent les tubules de l'épididyme qui se réunissent en un tubule de l'épididyme unique qui se jette dans le canal mésonéphrétique. Celui-ci devient le canal déférent. Le mésentère rattachant le testicule à la paroi abdominale prend le nom de mésorchium.
Chez la femelle, les cordons sexuels ne se creusent pas; les cellules originaires du cordon mésonéphrétique deviennent les cellules folliculaires entourant les cellules germinales ou ovogonies qui deviennent ovocytes. Une cellule germinale et les cellules folliculaires associées forment un follicule ovarien. Le mésentère est le mésovarium. La gonade mâle se développe un peu plus tôt et à un rythme plus rapide que la gonade femelle.
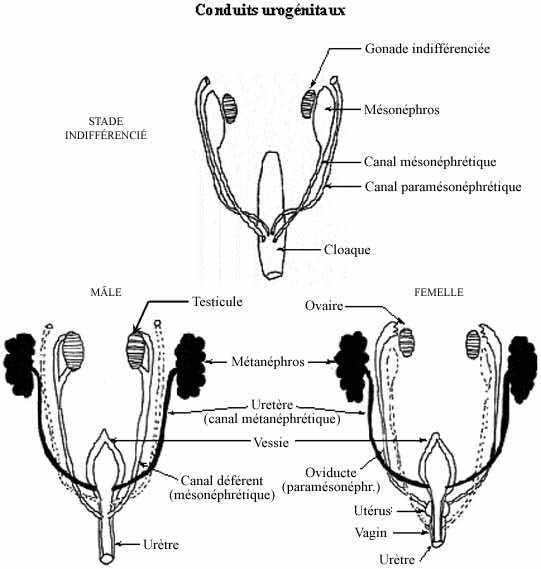
Chez les amphibiens, la médulla de l'ovaire dégénère et l'organe devient creux. L'ovulation se produit néanmoins à la face corticale de l'ovaire. Chez les oiseaux, seul l'ovaire gauche se développe complètement. La gonade droite demeure indifférenciée, avec des cordons cellulaires sans cellules germinales, celles-ci ayant dégénéré. Contrairement à la condition mâle, les canaux mésonéphrétiques dégénèrent chez l'embryon femelle. Une seconde paire de canaux se forme, les canaux paramésonéphrétiques (de Müller), tant chez le mâle que chez la femelle. Ils ne sont retenus que chez la femelle cependant: les oviductes, non continus avec les ovaires mais les recouvrant par leur extrémité agrandie en entonnoir. Ils dégénèrent chez le mâle.
Les deux canaux paramésonéphrétiques rencontrent postérieurement un renflement du cloaque, l'utérus, qui est en réalité la fusion des oviductes à leur extrémité caudale. Chez les sous-mammaliens, les oviductes demeurent séparés sur presque toute la longueur. Au niveau du cloaque, l'utérus rétréci devient le vagin, séparé par un septum de la portion urinaire du cloaque (l'urètre).
Chez le mâle, le canal mésonéphrétique, ou canal déférent, rejoint l'urètre passé la vessie, et l'urètre sert de canal excréteur tant pour l'urine que pour le sperme. Chez plusieurs mammifères, le canal déférent s'allonge considérablement, permettant la descente des testicules dans la paroi abdominale et même chez des espèces leur sortie extérieure dans un repli cutané périnéal appelé scrotum (dont l'homologue femelle constitue les lèvres majeures). Cette migration des testicules semble contrôlée par la testostérone et une autre substance sécrétée par les testicules. Chez certaines espèces scrotales, dont l'humain, les testicules sont en permanence dans le scrotum. Chez l'humain, les testicules atteignent le scrotum vers le 7e ou 8e mois de grossesse. Environ 1% des naissances humaines mâles se caractérisent par la condition appelée cryptorchidisme, i.e., un ou deux testicules ne se trouvent pas dans le scrotum (le taux est plus élevé chez les prématurés). Chez d'autres espèces scrotales, les testicules sont rétractables, migrant vers le scrotum à la période de reproduction et demeurant dans l'abdomen le reste du temps.
Voir schéma sur les conduits urogénitaux
- Détermination sexuelle chez les mammifères euthériens:
Le développement des mammifères se produit dans la direction "femelle", à moins d'être affecté par un produit mâle. Par exemple, en l'absence de gonades l'individu ressemble davantage à une "femelle", avec des voies ressemblant plutôt à celles de la femelle qu'à celles du mâle.
Le développement des testicules chez les mammifères semble dépendre de la présence du gène TDF/Tdy (aussi SRY) lié au chromosome Y, gène qui n'agirait pas seul mais en coopération avec un ou des gènes liés au chromosome X et aux autosomes. L'existence du gène TDF/Tdy déclencherait une cascade d'événements aboutissant à la formation des testicules tandis que son absence mènerait à une séquence d'événements différents résultant en la formation des ovaires. Les gènes autres que TDF/Tdy (i.e. ceux liés à X ou aux autosomes) sont les mêmes chez le mâle et la femelle. L'on ne sait pas encore combien de gènes au total sont impliqués dans la détermination sexuelle chez les mammifères.
Une fois formé, le testicule sécrète deux hormones principales:
testostérone: masculinise le foetus en stimulant la formation du pénis et autres structures, et en détruisant les primordia des glandes mammaires. Chez la femelle, la canal mésonéphrétique (de Wolff) privé de stimulation de testostérone régresse. Il s'agit donc d'une induction permissive chez le mâle.
AMH: hormone anti-müllerienne, une glycoprotéine qui détruit le canal paramésonéphrétique (de Müller), lequel se développe en oviducte chez la femelle.
Un gène analogue à TDF/Tdy aurait été conservé durant toute l'évolution des Vertébrés, chez le sexe hétérogamétique. Chez les mammifères, le mâle est XY (la femelle est XX); chez les oiseaux, la femelle est ZW (le mâle est ZZ).
Chez les mammifères marsupiaux, les caractères sexuels secondaires apparaissent avant la différenciation sexuelle des gonades; ils ne peuvent donc dépendre des hormones sexuelles produites par les gonades et seraient, semble-t-il, déterminés génétiquement.
ORGANOGENESE: DERIVES DU MESODERME 2:
SYSTEME CARDIOVASCULAIRE (cours 10)
On observe, chez les Métazoaires, une spécialisation des fonctions, et à ces fonctions diverses sont consacrés des organes divers. Une telle spécialisation morphologique et fonctionnelle nécessite un système de transport entre les différents organes. Chez la plupart des organismes, ce système de transport comprend un véhicule fluide amené d'un organe à l'autre par un réseau de conduits tubulaires grâce à une pompe qui inflige une pression au fluide.
Le fluide est de deux natures: le sang et la lymphe; les conduits de distribution sont les vaisseaux sanguins et lymphatiques; la pompe est le coeur pour le sang, la circulation lymphatique en est dépourvue.
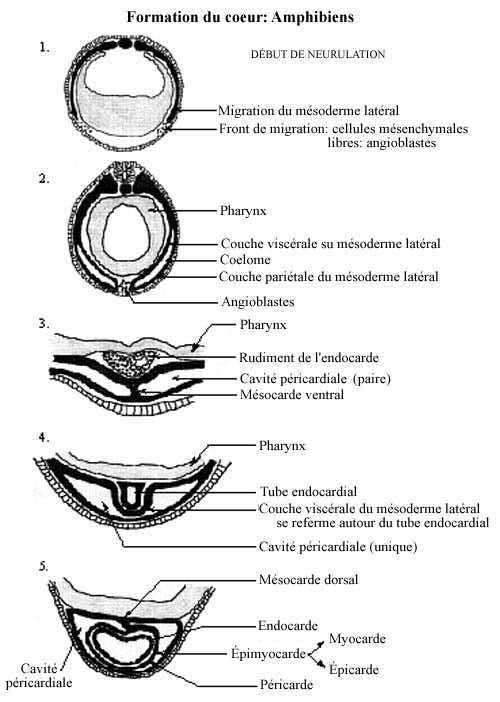
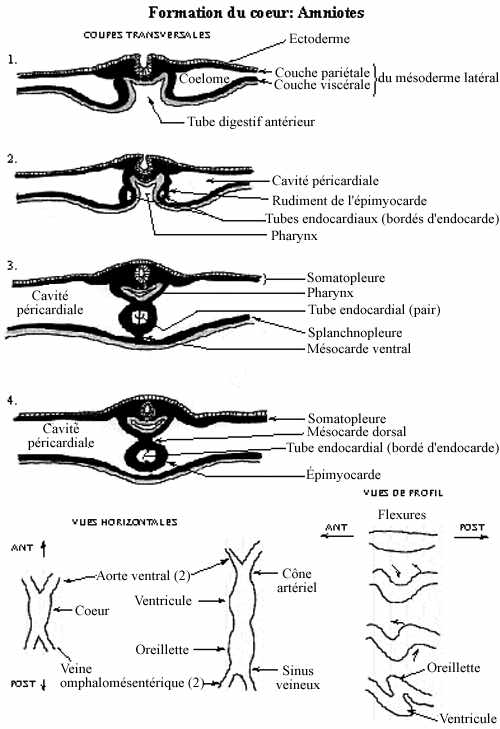
1- Formation du coeur
Au niveau du pharynx, des cellules mésodermiques se distinguent des autres cellules mésodermiques, et ceci dès l'étape de la neurulation. Ces cellules, libres, constituent les deux fronts de migration du mésoderme latéral qui progressent ventralement: ce sont les cellules angiogéniques ou angioblastes. Ces cellules mésodermiques déjà spécifiées ne se retrouvent qu'au niveau du pharynx, et non pas plus caudalement dans le mésoderme latéral.
Leur migration terminée, les angioblastes s'assemblent autour d'une cavité, formant le tube endocardial, détaché du reste du mésoderme latéral. Quant à ce dernier, les fronts de migration des côtés gauche et droit, en réalité les feuillets bilaminés décrits au chapitre précédent, se rejoignent ventralement au tube endocardial. En fusionnant, ils forment le mésocarde ventral, qui disparaîtra sous peu. Le tube endocardial repose donc sur le feuillet viscéral du mésoderme latéral fusionné. Le tube s'y enfonce, et le feuillet viscéral se referme autour de lui et y adhère: c'est l'épimyocarde. L'épimyocarde est relié dorsalement au reste du mésoderme latéral par le mésocarde dorsal, qui lui aussi est appelé à se résorber. Le coelome autour du coeur (entre les feuillets viscéral et pariétal du mésoderme latéral) s'appelle cavité péricardiale.
L'épimyocarde est à l'origine du myocarde, autour de l'endocarde, et de l'épicarde, autour du myocarde. Les cellules du myocarde se différencient en cellules musculaires cardiaques typiques et en cellules modifiées pour la conduction. Les cellules de l'épicarde se différencient, avec le matériel intercellulaire, en tissu conjonctif.
Le feuillet pariétal du mésoderme latéral renferme donc la cavité péricardiale: on appelle ce mésentère péricarde; il ne fait pas partie intégrante du coeur.
Au cours de la neurulation, des cellules du feuillet viscéral du mésoderme latéral, au niveau du pharynx, se différencient en angioblastes (cellules angiogéniques), non aisées à distinguer morphologiquement. Les angioblastes se retrouvent donc des côtés gauche et droit dans l'embryon, puisque le mésoderme latéral des deux côtés ne s'est pas encore rencontré ventralement. Au sein du groupe de cellules angiogéniques, bilatéralement, se creuse une cavité qui marque la formation du tube endocardial. Ce dernier est donc pair. Les cellules mésodermiques latérales au tube endocardial constituent le rudiment de l'épimyocarde. Le coelome qui borde latéralement ces structures cardiaques devient la cavité péricardiale, elle aussi paire.
Éventuellement, à mesure que se forme la tige vitelline, le mésoderme se referme ventromédialement. Au niveau du pharynx, ceci signifie l'apposition des coeurs gauche et droit: la formation d'un coeur unique divisé en moitiés gauche et droite par un septum d'endocarde. La fusion mésodermique ventromédiane amène la création d'un mésocarde ventral, qui se trouve à rattacher le coeur au reste du mésoderme latéral et qui disparaîtra rapidement, ainsi que d'un mésocarde dorsal qui rattache le coeur au mésoderme recouvrant le pharynx et qui disparaîtra également. Le filet d'endocarde qui divisait en deux la cavité cardiaque se désintègre, créant une chambre cardiaque unique.
Le coeur ainsi formé se trouve à "l'extérieur" de l'embryon. Ce n'est que plus tard qu'il migrera postérieurement et à l'intérieur de la cavité thoracique.
Chez les anamniotes comme chez les amniotes, les cellules de l'épimyocarde rudimentaire amorcent dès lors des mouvements de contraction rythmique, autonomes, avant même de recevoir une innervation. Celle-ci ne s'implante que plus tard. Le coeur est le premier organe fonctionnel de l'embryon.
La cavité péricardiale se sépare du reste du coelome par un repli mésodermique pariétal, le septum transverse, dans lequel se différenciera le diaphragme (muscle respiratoire), et le coeur devient ainsi isolé des organes rudimentaires de la cavité abdominale.
À chacune de ses extrémités, le coeur bifurque. Antérieurement émergent les deux aortes ventrales, postérieurement les deux veines omphalomésentériques. Au point d'émergence des aortes ventrales, à partir du ventricule, se trouve le cône artériel, et au point d'émergence des veines omphalomésentériques, partant de l'oreillette, se trouve le sinus veineux.
Une constriction délimite deux dilatations du coeur: le ventricule antérieurement (se continuant dans le cône artériel) et l'oreillette postérieurement (se continuant dans le sinus veineux). Initialement droit, le coeur subit des flexures, un peu comme le tube neural céphalique. Leur ampleur varie chez les différents groupes mais chez tous on en retrouve un minimum. Ainsi, le bout postérieur du coeur est repoussé vers l'avant, moyennant une pliure vers le bas, puis une vers la droite. D'autres replis se produisent vers la gauche, le haut et l'avant. La portion du coeur autrefois postérieure, l'oreillette, devient maintenant en position dorsale et antérieure. La portion originalement antérieure, le ventricule, se retrouve ventrale et postérieure. Les repliements du coeur obéissent à des forces intrinsèques. Un résultat de ces mouvements est que le coeur est repoussé vers la moitié gauche de l'animal. Ceci est probablement le premier indice de latéralisation d'un organe, une notion amenée à la fin du chapitre sur la gastrulation. D'autres organes se latéralisent plus tard.
Chez les Vertébrés pulmonés, donc développant une circulation pulmonaire, le coeur se sépare en deux moitiés. La moitié droite reçoit le sang désoxygéné des différents organes (par les veines caves) et apporte ce sang aux poumons (par les artères pulmonaires); la moitié gauche reçoit le sang oxygéné des poumons (par les veines pulmonaires) et l'envoie aux autres organes de l'animal (par l'aorte).
Cette séparation s'effectue à des degrés plus ou moins avancés, selon les groupes zoologiques. Elle s'annonce déjà chez les poissons dipneustes. Chez les amphibiens, une partition s'établit dans le sinus veineux et l'oreillette. L'oreillette droite est rattachée aux veines caves, l'oreillette gauche à la veine pulmonaire qui bifurque immédiatement vers les poumons. Les oreillettes déversent cependant leur sang dans un ventricule unique. Chez les amniotes, oreillettes et ventricules sont pairs, sauf pour certains reptiles qui présentent des stades intermédiaires.
2- Formation des vaisseaux sanguins (voir schéma)
Il semble que chaque portion de l'organisme qui nécessite un apport sanguin soit effectivement pourvue en vaisseaux sanguins. Les artères et veines se forment in situ à partir de cellules du mésoderme local appelées angioblastes. Les cellules angioblastiques peuvent provenir de toutes les régions mésodermiques de l'embryon, sauf la notochorde et la plaque préchordale. Ceci inclut donc les somites, le mésoderme intermédiaire, les deux feuillets du mésoderme latéral et le mésenchyme interstitiel. Les angioblastes peuvent migrer sur des distances considérables, avant même de s'assembler en tubules (voir ci-après), leurs directions de migration n'étant altérées que par le contact avec les tissus épithéliaux. Les vaisseaux de la tête ne se forment pas à partir du mésenchyme originaire de la crête neurale mais uniquement à partir de mésoderme véritable.
Les angioblastes s'assemblent en un épithélium simple pavimenteux, l'endothélium, autour d'une lumière, formant une structure tubulaire, les tubules endothéliaux, à diamètre constant. Le signal permettant aux angioblastes de s'assembler en tubules, déterminant donc le site de formation des vaisseaux, provient d'interactions avec l'environnement local et est indépendant de l'origine des angioblastes.
Les vaisseaux originaux ne sont pas droits mais tortueux et forment des réseaux, des anastomoses. Une fois le réseau de tubules endothéliaux en place, des tubules peuvent se former par embranchement de tubules existants, de nouveaux tubules peuvent se connecter aux tubules en place, des tubules peuvent disparaître; tout cela selon les besoins locaux. Donc, une plasticité considérable se manifeste dans la formation des vaisseaux sanguins d'une région donnée; il en découle une certaine variation d'un individu à l'autre.
Le développement subséquent de la paroi des vaisseaux dépend de la quantité et de la direction du sang qui y circule. Les tubules recevant plus de sang ou du sang sous plus forte pression ajoutent à l'épaisseur de leur paroi. Autour de l'endothélium s'additionnent muscle lisse et tissu conjonctif à partir du mésoderme local. La paroi des vaisseaux renferme trois couches, plus ou moins bien développées selon le type de vaisseau (artère ou veine) et selon le diamètre du vaisseau:
- tunica intima: endothélium et tissu conjonctif délicat; seule couche des capillaires- tunica media: muscle lisse et tissu conjonctif élastique
- tunica adventitia: tissu conjonctif
Le coeur et les vaisseaux sont formés avant que ne s'établisse la circulation du sang.
3- Formation des cellules sanguines
Les cellules sanguines se différencient à partir de cellules souches pluripotentes: "colony forming units" ou CFU. Chez les mammifères, seul le sac vitellin serait à l'origine des CFU? Chez les oiseaux et autres, le sac vitellin et des îlots sanguins locaux dans des tubules endothéliaux seraient à l'origine de CFU.
À partir du sac vitellin, des CFU migrent d'abord au foie en formation, puis à la rate en formation, puis à la moelle osseuse, où s'effectue à tour de rôle l'hémopoïèse chez l'embryon. Chez plusieurs animaux mûrs, seule la moelle de certains os, et parfois la rate, retiennent la fonction d'hémopoïèse. Ce qui induit les CFU à se différencier en cellules sanguines semble venir de l'environnement local: "hemopoietic inductive microenvironment" ou HIM.
4- Circulation sanguine
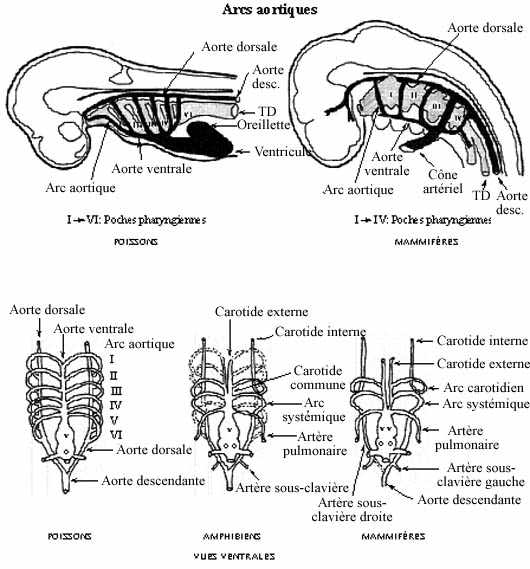
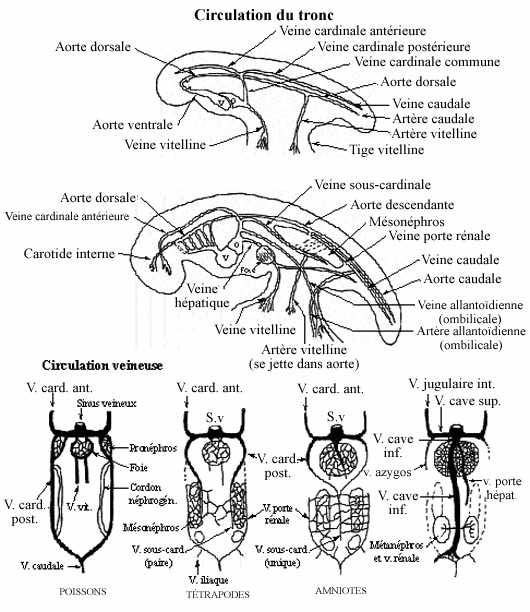
- Circulation artérielle (voir schéma)
Les aortes ventrales, continuations du cône artériel, amènent le sang rostralement, vers le pharynx et la tête. Une paire d'aortes dorsales se développe in situ dorsalement au pharynx; l'aorte dorsale est connectée à l'aorte ventrale du même côté par 6 arcs aortiques qui traversent les arcs pharyngiens (branchiaux), dont nous avons déjà parlés. Les arcs aortiques se développent à partir du mésenchyme local selon un gradient antéro-postérieur. Six paires d'arcs aortiques se développent, même chez les Vertébrés qui ne possèdent que 4 paires d'arcs pharyngiens. Les deux aortes dorsales se réunissent postérieurement au pharynx pour former l'aorte descendante qui parcourt le tronc, dorsalement au tube digestif. Chez les Vertébrés aquatiques (poissons et amphibiens larvaires), les arcs aortiques 2 à 5 apportent le sang aux branchies, organes de la respiration. À l'intérieur des branchies se développent des réseaux capillaires. Le retour sanguin des branchies s'effectue via les aortes dorsales, continues en aorte descendante, qui amènent le sang oxygéné au corps. Les réseaux capillaires (en fait, les branchies) sont perdus chez les amphibiens qui se métamorphosent mais un vaisseau continue de relier les aortes ventrales et dorsales aux mêmes endroits.
Les 2 premières paires d'arcs aortiques dégénèrent plus ou moins chez les amphibiens et totalement chez les amniotes.
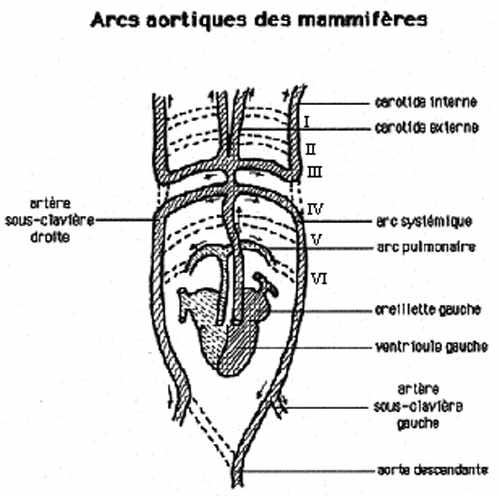
La 3e paire devient une paire d'arcs carotidiens. Rostralement aux arcs carotidiens, les deux aortes ventrales deviennent les artères carotides externes et les aortes dorsales deviennent les artères carotides internes. L'arc carotidien est relié au 4e arc aortique par l'artère carotide commune, de chaque côté.
La 4e paire d'arcs aortiques devient une paire d'arcs systémiques. Entre les arcs carotidiens et systémiques les aortes dorsales dégénèrent.
La 5e paire d'arcs aortiques dégénère.
La 6e paire devient une paire d'artères pulmonaires, qui perdent leur connexion avec la paire d'aortes dorsales.
Juste avant la jonction des aortes dorsales pour former l'aorte descendante, émerge une paire d'artères sous-clavières, desservant les membres antérieurs notamment.
Chez les mammifères, l'arc systémique gauche demeure tel que décrit chez les autres amniotes mais le droit perd sa connexion avec l'aorte descendante et devient l'artère sous-clavière droite. L'artère sous-clavière gauche émerge toujours de l'aorte dorsale gauche, maintenant déconnectée de la droite.
La réduction et la disparition de certains arcs aortiques et portions de vaisseaux dépendent de la diminution du flot sanguin qui les parcourt.
À partir de l'aorte descendante émergent des artères (paires) irriguant chaque métamère (les artères intersegmentaires), des artères irriguant les viscères, une paire d'artères irriguant les membres postérieurs (les 2 artères iliaques), en plus, chez les amniotes, d'artères irriguant le sac vitellin (les 2 artères vitellines) et l'allantoïde (les 2 artères allantoïdiennes). Par analogie, on donne souvent le nom d'artères vitellines aux artères irriguant la paroi du tube digestif chez les anamniotes. L'aorte descendante se poursuit jusqu'à la queue (artère caudale).
- Circulation veineuse (voir schéma)
Les veines cardinales antérieures drainent le sang de la tête et de la région pharyngienne, les veines cardinales postérieures le sang du tronc, dont du pronéphros. Ces dernières sont le résultat de la bifurcation de la veine caudale unique qui draine la queue. Les deux paires de veines cardinales se jettent dans les veines cardinales communes reliées au sinus veineux.
Une paire de veines vitellines apporte le sang enrichi des produits nutritifs, du sac vitellin (paroi du tube digestif chez les anamniotes) au sinus veineux. Elles passent au travers du foie en formation où elles établissent un réseau capillaire, les sinusoïdes hépatiques. La portion des veines vitellines entre le foie et le sinus veineux prend le nom de veines hépatiques.
Quand se développe le mésonéphros, une paire de veines sous-cardinales s'y forme médialement.
Comme les pronéphros disparaissent, la portion des veines cardinales postérieures qui les irriguait dégénère; la portion postérieure au bris devient veine porte rénale, latérale au mésonéphros, la portion antérieure au bris se connecte à la veine sous-cardinale. Les veines sous-cardinales, médiales aux mésonéphros, sont connectées aux veines portes rénales, latérales aux mésonéphros, par des réseaux capillaires traversant les mésonéphros.
Chez les tétrapodes, une paire de veines iliaques, se jetant dans les veines portes rénales, dessert les membres postérieurs.
Chez les amniotes, à mesure que disparaissent les mésonéphros, les deux veines sous-cardinales migrent médialement et fusionnent pour former une veine sous-cardinale unique, médiane. La veine vitelline droite s'y connecte, le tout devenant veine cave inférieure. La veine vitelline gauche devient veine porte hépatique, qui draine les intestins.
Les veines portes rénales disparaissent. Dans la veine cave inférieure se jettent une paire de veines rénales, associées aux métanéphros, et la paire de veines iliaques. Les veines cardinales postérieures disparaissent, sauf une portion de la droite qui devient veine azygos. Les veines cardinales antérieures deviennent veines jugulaires internes, les veines cardinales communes deviennent veine cave supérieure, qui reçoit la veine azygos.
Chez les amniotes les veines allantoïdiennes (ombilicales chez les mammifères) atteignent le sinus veineux via les veines cardinales communes.
Chez le foetus mammifère, une ouverture dans la paroi séparant l'aorte et l'artère pulmonaire, le ductus arterosius, dirige le sang de l'artère pulmonaire vers l'aorte et, donc, vers le placenta. Il n'y a pas de sang acheminé vers les poumons par l'artère pulmonaire, ni de sang qui en revienne par la veine pulmonaire dans l'oreillette gauche. Pour que le sang rejeté dans l'oreillette droite soit pompé dans la circulation systémique, une ouverture dans la paroi interauriculaire, le foramen ovale, le laisse s'échapper vers l'oreillette gauche et, de là, au ventricule gauche.
À la naissance, le nouveau-né n'est plus rattaché au placenta et le sang doit être immédiatement redistribué vers les poumons. À la première respiration, l'O2 sanguin élevé induit les muscles septaux entourant le ductus arteriosus à le refermer, séparant ainsi complètement aorte et artère pulmonaire. Comme la pression du côté gauche du coeur augmente, elle fait refermer une valve qui clôt le foramen ovale, séparant complètement oreillettes gauche et droite, et donc les circulations pulmonaire et systémique. Quand la respiration s'amorce chez le nouveau-né, la circulation respiratoire est dirigée du placenta vers les poumons.
Croissance
Jusqu'à présent nous avons étudié les deux premières étapes de la phase progressive du cycle vital. Une fois les rudiments des organes en place, le développement subséquent se caractérise non pas tant par des changements ou des remaniements de tissus comme par l'accroissement de la taille des structures en place. Ainsi, une durée importante du développement animal consiste en une étape de croissance. Chez le mammifère, cette période de croissance suivant le développement embryonnaire s'appelle période foetale, et l'embryon prend le nom de foetus.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}