SEGMENTATION OU CLIVAGE (cours 4)
Chez toutes les espèces connues, la production d'un organisme pluricellulaire commence par une étape appelée segmentation, qui consiste en une série de divisions mitotiques. L'oeuf fécondé, plus ou moins considérable selon les groupes zoologiques, est segmenté en de nombreuses petites cellules appelées blastomères. La masse de cellules résultante s'appelle blastula et est généralement creusée d'une cavité, le blastocoele, remplie de fluide. La segmentation est le résultat de deux processus coordonnés:caryocinèse: division mitotique du noyau, dont l'agent mécanique est le fuseau mitotique avec ses microtubules composés de tubuline.
cytocinèse: division de la cellule entière, dont l'agent mécanique est l'anneau contractile de microfilaments composés d'actine.
Chez la plupart des espèces il n'y a pas de gain volumétrique total durant la segmentation. Ceci résulte de l'absence de période de croissance interphasique (entre les divisions) qui caractérise normalement le cycle cellulaire. Durant la segmentation, la cytocinèse s'effectue à un rythme accéléré tandis qu'aucun cytoplasme nouveau est ajouté aux cellules-filles: la masse cytoplasmique de l'oeuf leur est répartie. Ainsi les cellules-filles deviennent de plus en plus petites au fur et à mesure que progresse la segmentation.
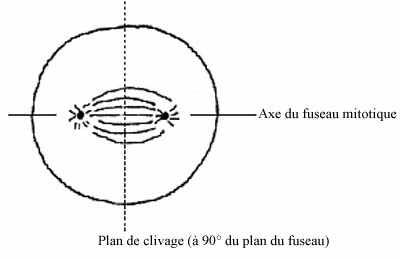
Les asters du fuseau mitotique dictent le plan de division cellulaire: ce dernier est toujours perpendiculaire à l'axe du fuseau mitotique. Le premier plan de clivage est spécifié par le point d'entrée du spermatozoïde dans l'oeuf, qui amène une réorganisation du cytoplasme de l'oeuf, notamment du cytosquelette, incluant une protéine importante de celui-ci, la vimentine. Donc, le centriole mâle agit comme centre organisateur.
Outre cette influence directe du centre organisateur mâle sur le premier plan mitotique du zygote, d'autres facteurs agissent sur la segmentation:
1) des facteurs dans le cytoplasme (morphogènes) de l'oeuf peuvent influencer le fuseau mitotique, dont le moment de sa formation;
2) la quantité et la localisation du vitellus dans le cytoplasme. Le vitellus détermine où peuvent s'effectuer les divisions, leur cinétique, la taille relative des blastomères et du blastocoele ainsi que la durée totale de la période de segmentation. Les divisions se déroulent plus rapidement au pôle pauvre en vitellus, le pôle animal, qu'au pôle riche en vitellus, le pôle végétatif. Ceci sera clarifié bientôt.
Tout en tenant compte de l'influence du centre organisateur mâle sur le premier plan de clivage, le type de segmentation, c'est-à-dire la séquence des plans de division, des cinétiques mitotiques et la grosseur relative des blastomères, est caractéristique à un groupe zoologique et est invariable (sous contrôle génétique).
Les oeufs ne contenant pas une grande quantité de vitellus subissent une segmentation holoblastique: les plans de clivage traversent l'oeuf entier. Selon l'angle de ses axes, elle peut être:
radiaire
spiralée
bilatérale
rotationnelle
Les oeufs renfermant beaucoup de vitellus subissent une segmentation méroblastique: seulement une portion du cytoplasme se segmente car l'axe de division ne pénètre pas ou ne pénètre qu'incomplètement le vitellus. La segmentation méroblastique peut être:
discoïde chez les oeufs télolécithes: vitellus abondant au pôle végétatif
superficielle chez les oeufs centrolécithes: vitellus abondant central
1) Clivage holoblastique radiaire:
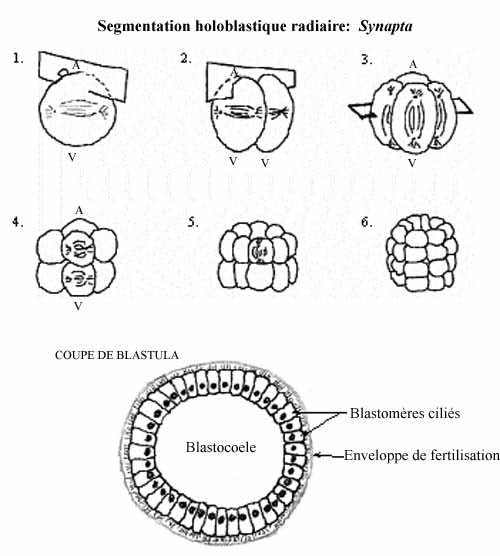
Ex.1: le concombre de mer Synapta, un échinoderme:
Le concombre de mer présente la forme la plus simple de segmentation. L'oeuf des échinodermes est oligolécithe, il renferme peu de réserves vitellines et celles-ci sont réparties uniformément dans le cytoplasme. Après l'union des deux pronoyaux, l'axe du premier fuseau mitotique se forme perpendiculaire à l'axe A-V de l'oeuf fécondé; donc le premier clivage passe directement au travers des pôles animal et végétatif. Comme le plan de clivage coupe ces deux pôles, on dit qu'il est méridional. Comme les deux blastomères obtenus sont de taille égale, c'est un clivage égal.
Les fuseaux mitotiques de la seconde division sont également perpendiculaires à l'axe A-V, mais à angle droit par rapport au premier fuseau. La division coupe les pôles A-V, se produit simultanément dans les deux blastomères et résulte en la formation de 4 blastomères égaux. Elle est méridionale et égale.
Le troisième clivage ne passe plus par les pôles A-V de l'embryon mais, pour chacun des 4 blastomères, est perpendiculaire à cet axe car les fuseaux mitotiques sont dans l'axe A-V. On dit que le clivage est équatorial. Il sépare l'embryon en 8 blastomères égaux.
La quatrième division est méridionale et produit 16 blastomères égaux.
Les divisions successives se font en alternance selon les plans équatorial et méridional, jusqu'au stade 256 cellules.
Le résultat de la segmentation est une petite boule, la blastula, dont le volume est sensiblement égal à celui de l'oeuf initial et dont les petits blastomères ont tous la même taille. Au cours des divisions, les contacts intercellulaires se sont relâchés de sorte que les blastomères ne s'agrègent pas en une masse compacte, mais décrivent une sphère d'une seule assise cellulaire autour d'une cavité remplie de fluide, le blastocoele. Les cellules qui se jouxtent établissent des jonctions entre elles. La blastula est entourée de l'enveloppe de fertilisation.
Les blastomères sécrètent des protéines dans le blastocoele; le fluide qu'il renferme devient sirupeux et a la propension de retenir beaucoup d'eau. Ainsi, au cours de la segmentation, de l'eau pénètre dans le blastocoele par osmose, à partir du milieu aqueux externe, et fait enfler la blastula. Cette pression exercée sur les blastomères pourrait aider à les orienter de sorte à ce qu'ils décrivent une sphère d'une seule assise cellulaire autour du blastocoele.
Les cellules possèdent des cils à leur surface externe. Leur battement impute à la blastula un mouvement de rotation à l'intérieur de l'enveloppe de fertilisation. Peu après la fin de la segmentation, les blastomères sécrètent une enzyme, l'enzyme d'éclosion, qui digère l'enveloppe de fertilisation. L'embryon devient une larve nageant librement.
À n'importe quel moment pendant la segmentation, un embryon coupé en deux selon n'importe quel méridien produira deux moitiés qui sont identiques. Ce type de symétrie est caractéristique d'une sphère ou d'un cylindre, il s'agit de la symétrie radiaire.
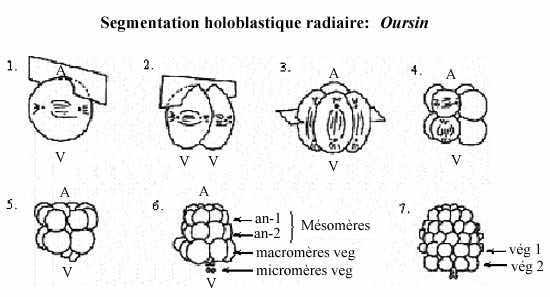
Ex.2: l'oursin, un échinoderme
L'oursin possède lui aussi un oeuf oligolécithe.
Ses 3 premiers clivages ressemblent à ceux du concombre de mer Synapta. Au 4e clivage, les 4 blastomères de l'hémisphère animal se divisent selon l'axe méridional et donnent une assise de 8 blastomères de taille égale appelés mésomères; les blastomères de l'hémisphère végétatif se divisent selon l'axe équatorial mais inégalement, en réalité sous l'équateur, et donnent 4 grosses cellules appelées macromères, sous les mésomères, et 4 petits micromères, sous les macromères.
Au clivage suivant, les 8 mésomères se divisent équatorialement (également), donnant deux assises animales, l'une au dessus de l'autre: an-1 et an-2. Les macromères se divisent méridionalement, donnant une assise de 8 macromères; les micromères, en se divisant, forment un petit amas sous les macromères.
Tous les plans de clivage de la 6e division sont équatoriaux; les macromères forment donc deux assises végétatives, l'une au dessus de l'autre: vég-1 et vég-2. Les 7e divisions sont méridionales. Il en résulte une blastula de 128 cellules, creusée d'un blastocoele rempli de fluide, et entourée de l'enveloppe de fertilisation. Une larve se forme comme chez Synapta.
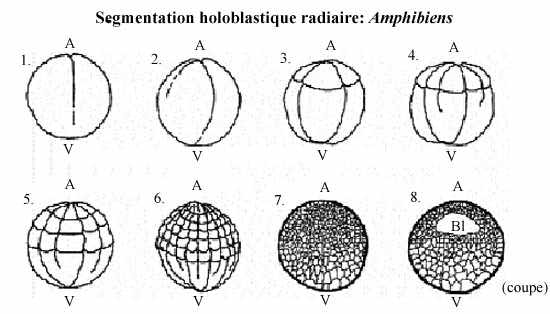
Ex.3: les amphibiens
L'oeuf mésolécithe d'amphibien possède plus de vitellus que celui des échinodermes, vitellus qui devient concentré au pôle végétatif après la fécondation (oeuf hétérolécithe), et le noyau migre au pôle animal.
La 1ère division commence au pôle animal de l'oeuf et s'étend lentement vers le pôle végétatif; elle est méridionale et égale. Avant que cette division ne soit achevée, un second clivage méridional s'amorce au pôle animal, perpendiculaire au premier, lui aussi égal.
Le 3e clivage est équatorial; il ne s'effectue pas au centre de l'embryon mais plus près du pôle animal (donc, sus-équatorial), là où les plans de clivages des deux divisions précédentes ont achevé de traverser; il est donc inégal. Les plans de clivage méridionaux, du moins le second, n'ont pas encore achevé de traverser le cytoplasme de l'hémisphère végétatif quand a lieu ce 3e clivage sus-équatorial. Cette division sépare l'embryon en 4 petits blastomères animaux, les micromères, et 4 gros macromères végétatifs (qui éventuellement s'individualisent). Ce clivage holoblastique inégal établit deux régions embryonnaires majeures: une région de micromères qui se diviseront rapidement, dans l'hémisphère animal, et une région de macromères qui se diviseront lentement, dans l'hémisphère végétatif.
Les embryons amphibiens aux stades 16 à 64 cellules portent le nom de morula, à cause de leur aspect de mûre. Au stade 128 cellules l'embryon est considéré une blastula, creusée d'un blastocoele de volume proportionnellement moindre que celui des échinodermes. Il occupe l'hémisphère animal; son plafond est constitué de plus d'une couche de micromères et son plancher de plusieurs assises de macromères.
Le blastocoele est une cavité qui permettra l'occurrence des migrations cellulaires pendant la gastrulation. Il empêche une interaction prématurée entre les cellules qu'il sépare. Par exemple, si les cellules du toit du blastocoele étaient apposées à celles du plancher, au lieu de se différencier en ectoderme comme elles le font normalement, elles se différencieraient en mésoderme. Les cellules du plancher du blastocoele, qui se différencieront en endoderme, induisent la différenciation des cellules qui leur sont voisines en mésoderme. Nous reviendrons sur ces notions. Donc, le blastocoele permet aux cellules de la blastula de conserver leur identité.
2) Clivage holoblastique spiralé (voir schéma):
Se rencontre notamment chez les mollusques bivalves et les annélides, qui ont des oeufs isolécithes, pauvres en vitellus.
Au lieu de se produire parallèle ou perpendiculaire à l'axe A-V de l'oeuf, le clivage se produit à angle oblique.
En général, les deux premières divisions sont à peu près méridionales et égales.
À chaque clivage subséquent le plan de segmentation est oblique à l'axe A-V et perpendiculaire au plan de segmentation précédent. Chaque blastomère produit deux cellules de tailles différentes: un micromère dans l'hémisphère animal et un macromère dans l'hémisphère végétatif. Pour que les micromères et les macromères aient cette localisation préférentielle, ils migrent vers leur hémisphère respectif après la division. C'est ce mécanisme qui crée la relation en spirale caractéristique de ce type de clivage.
L'orientation du plan de clivage à gauche: spirale senestre (sens contraire aux aiguilles d'une montre), ou à droite: spirale dextre (dans le sens des aiguilles d'une montre), est contrôlée par des facteurs cytoplasmiques et déterminée génétiquement.
Cette blastula ne renferme pas de blastocoele et porte le nom de stéréoblastula. Ses cellules sont en contact sur une plus grande surface que ne le sont les blastomères qui subissent un clivage radiaire et qui abritent un blastocoele. Ces embryons accomplissent un moins grand nombre de divisions avant d'entreprendre l'étape subséquente de développement.
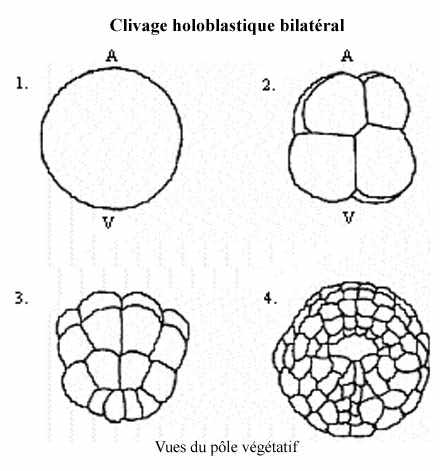
3) Clivage holoblastique bilatéral (voir schéma):
Se rencontre chez les ascidiens qui possèdent des oeufs oligolécithes, pauvres en vitellus.
Le 1er plan de segmentation, méridional, établit le seul plan de symétrie de l'embryon. Chaque division successive s'oriente selon ce plan de symétrie; le demi-embryon d'un côté de ce plan (et seulement de ce plan) est une image-miroir de l'autre.
Le 2e clivage est méridional mais ne passe pas par le centre de l'embryon. Il crée 2 gros blastomères antérieurs et 2 petits blastomères postérieurs. Ainsi, chaque moitié (établie par le premier clivage) possède un gros et un petit blastomère.
Aux 3 divisions suivantes, les différences de taille et de forme cellulaires accentuent la symétrie bilatérale de l'embryon. Au stade 32 cellules, un blastocoele est formé et la gastrulation commence.
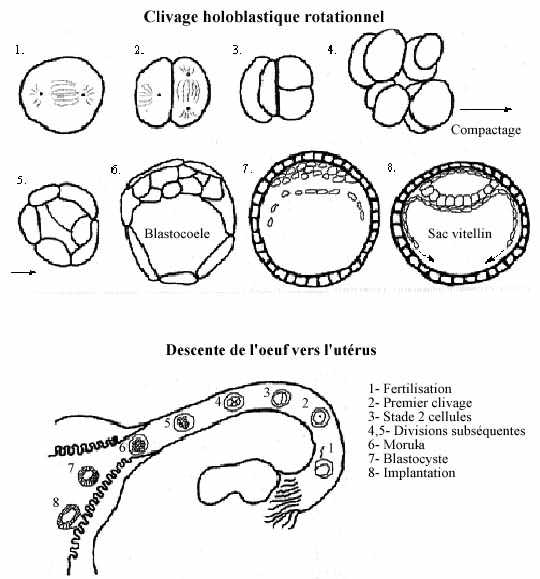
4) Clivage holoblastique rotationnel (voir schéma):
Se retrouve chez les mammifères dont les oeufs appartiennent au type alécithe.
La segmentation est tardive et lente chez les mammifères. Ceci est relié au fait que l'ovocyte a accumulé peu de réserves durant sa phase d'accroissement. Le premier clivage peut ne se produire qu'un jour après la fertilisation et les divisions peuvent être espacées de 12 à 24 heures, allouant une certaine interphase qui permet la transcription du génome embryonnaire. La 1ère division est méridionale et égale. La seconde diffère de tous les patrons déjà vus: un des deux blastomères se divise méridionalement, l'autre équatorialement; c'est le clivage rotationnel. De plus, les deux mitoses du second clivage peuvent ne pas se produire en synchronie, permettant des stades 3, 5, 6 cellules.
La segmentation des mammifères présente une autre singularité, le phénomène de compactage. Suivant le troisième clivage (stade 8 cellules), les blastomères maximisent les surfaces de contact entre eux. Quelques uns adoptent une forme généralement arrondie, en boule compacte: c'est la masse cellulaire interne ou bouton embryonnaire. D'autres blastomères adoptent une forme pavimenteuse et entourent la MCI; ils sont reliés entre eux par des jonctions cellulaires serrées: c'est le trophoblaste ou trophectoderme.
Un blastocoele se forme sous la MCI, délimité de l'autre part par une portion du trophoblaste. L'embryon entier prend le nom de blastocyste.
Le trophoblaste ne produira pas de structures embryonnaires proprement dites mais formera une portion externe du placenta. La MCI formera l'embryon ainsi que des structures extra-embryonnaires. Les cellules de la MCI diffèrent morphologiquement, fonctionnellement et biochimiquement de celles du trophoblaste. Néanmoins, le destin de chacun de ces deux types de cellules peut être inversé pendant toute la période de segmentation en faisant occuper à une la place de l'autre. L'appartenance des cellules à la MCI ou au trophoblaste dépend de leur position relative dans le blastocyste. Nous reviendrons sur ces notions au dernier chapitre.
La création de la MCI constitue le phénomène crucial des premiers stades du développement mammalien. Une fois le trophoblaste formé, l'embryon est capable de s'implanter dans la paroi utérine.
Des cellules se séparent de la MCI et forment l'hypoblaste qui tapisse le blastocoele et donnera naissance à l'endoderme du sac vitellin, une structure extra-embryonnaire. Ce qui reste de la MCI, au dessus de l'hypoblaste, est l'épiblaste. Ses cellules ne sont pas reliées par des jonctions serrées et l'épiblaste se délamine: la couche profonde produit l'épiblaste embryonnaire, et la couche supérieure produit la paroi de l'amnion, une autre structure extra-embryonnaire. Entre l'épiblaste embryonnaire et la paroi amniotique, la cavité amniotique est remplie du fluide amniotique; elle sert d'amortisseur et empèche la dessication de l'embryon.
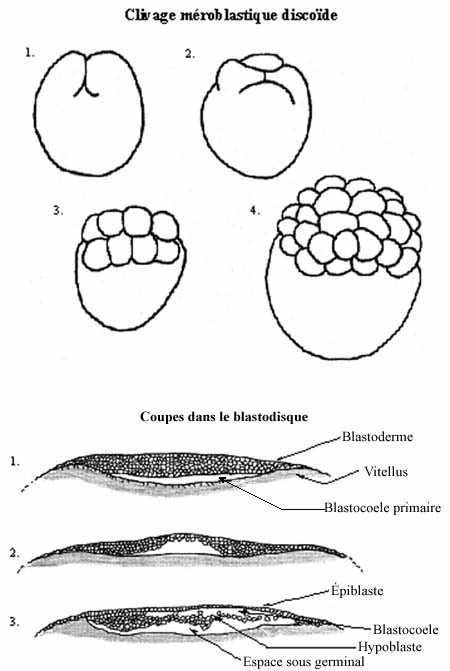
5) Clivage méroblastique discoïdal (voir schéma):
Il se retrouve chez les oiseaux, les reptiles et plusieurs poissons, dont les oeufs accumulent une grande quantité de vitellus.
La plus grande portion de l'oeuf télolécithe étant occupée par le vitellus, le noyau et le cytoplasme actif sont repoussés au pôle animal, seul affecté par la segmentation. Les 8 premiers blastomères, résultats de divisions méridionales, forment une assise sur la masse vitelline; leur membrane cytoplasmique est continue avec celle entourant le vitellus et leur cytoplasme repose sur le vitellus.
Des clivages équatoriaux amènent la formation de quelques assises de cellules complètement délimitées, mais l'assise inférieure repose toujours sur le vitellus. Les blastomères complets forment un disque à la surface de la masse vitelline, c'est la discoblastula ou disque germinal (non pas dans le sens de sexuel mais dans celui d'embryon). Sous le disque de blastomères complets se crée un espace, la cavité sous-germinale ou blastocoele primaire. L'ensemble des blastomères complets prend le nom de blastoderme. La structure entière constitue la blastula primaire.
Vu de surface le disque blastodermique se distingue en deux régions: l'aire pellucide, au centre du disque et qui se trouve au dessus du blastocoele primaire, d'où son apparence claire, et l'aire opaque, à la marge de l'aire précédente et qui repose sur le vitellus.
Des cellules du blastoderme se détachent et sont rejetées dans le blastocoele primaire, formant une lâche couche appelée hypoblaste; la couche supérieure constitue l'épiblaste. Le blastocoele primaire se trouve maintenant divisé en deux par l'hypoblaste, et la cavité entre ce dernier et l'épiblaste est le blastocoele, homologue à celui des amphibiens. Épiblaste et hypoblaste sont apposés tout autour du blastocoele; c'est ce qui constitue l'aire opaque. Aucune cellule de l'hypoblaste ne contribue à la formation de l'embryon. Il formera une partie des structures extra-embryonnaires. L'épiblaste formera une autre partie des structures externes ainsi que les tissus de l'embryon. C'est le stade qu'a atteint l'embryon lorsqu'a lieu la ponte de l'oeuf.
6) Clivage superficiel:
Rencontré chez les oeufs centrolécithes d'insectes, riches en vitellus central.
La segmentation offre ici aussi un exemple unique. Le noyau du zygote, en plein dans la masse vitelline, entreprend de nombreuses caryocinèses sans que ne se produisent de cytocinèses; ces noyaux s'appellent énergides. Ils migrent à la périphérie de l'oeuf, et les caryocinèses s'y poursuivent à un rythme prodigieux. L'embryon porte le nom de blastoderme syncitial: tous les noyaux du clivage sont contenus dans un même cytoplasme.
Des noyaux migrent au pôle postérieur de l'oeuf et s'enveloppent de membrane cytoplasmique; ce sont les cellules polaires de l'embryon. Elles donneront naissance aux cellules germinales (reproductrices) de l'animal adulte. Ainsi, un des premiers événements de l'embryogenèse des insectes est la distinction des cellules germinales et somatiques.
Après la formation des cellules polaires, la membrane ovocytaire s'invagine entre les noyaux, formant plusieurs petites cellules et créant le blastoderme cellulaire. Les cellules forment une seule assise autour du vitellus central. Chez la drosophile, par exemple, cette couche comprend jusqu'à 5000 cellules et est formée en dedans de 3 heures. Les cellules ne sont pas distribuées également autour du vitellus, ni au hasard. Il en existe une grande concentration à la face ventrale: c'est l'anlage germinal qui produira toutes les cellules de l'embryon. Les autres cellules de la couche unique formeront les structures extra-embryonnaires.
Après avoir comparé ces différents types de segmentation, on réalise que le vitellus est une adaptation évolutive permettant à l'embryon de se développer en l'absence de source externe de nourriture. Les animaux dont l'oeuf est pauvre en réserves vitellines développent généralement un stade larvaire assez rapidement (dès la fin du clivage chez l'oursin); la larve peut se nourrir et nager librement, alors que le développement continue.
Les oeufs des mammifères placentaires ne possèdent pas de vitellus mais leur stratégie diffère. Ils développent un placenta relié à la mère, qui leur procure nourriture et oxygène. D'ailleurs, une des premières différenciations de l'embryon mammalien est celle des cellules servant à former le placenta.
À l'autre extrême, les oeufs des oiseaux, des reptiles et de certains poissons contiennent des réserves vitellines suffisantes pour que les embryons puissent se nourrir sans avoir recours à un stade larvaire ni à un placenta.
La segmentation débute peu après la fécondation et se poursuit plus ou moins rapidement et plus ou moins longtemps, selon les espèces, jusqu'à ce qu'une quelconque balance noyaux/cytoplasmes soit atteinte. Par exemple, l'oeuf de la grenouille Rana pipiens produit 37 000 blastomères en 43 heures, à 15°C.
Donc, chez la plupart des groupes zoologiques, un bon nombre de clivages se produit avant que la transcription de nouveaux messagers par les blastomères ne soit en branle. Par exemple, chez la grenouille Xenopus laevis la transcription d'ARNm du génome embryonnaire n'est pas activée avant la 12e division. C'est donc dire que le matériel requis pour les 12 premières divisions a été fourni à partir des réserves d'ARNm et de protéines maternels, accumulés pendant l'ovogenèse et transmis au zygote. Les mitochondries dérivent aussi exclusivement du stock maternel.
Plus les réserves (autres que vitellines) accumulées durant l'ovogenèse et transmises au zygote sont considérables, plus il pourra se produire un nombre élevé de clivages rapides sans avoir recours au génome embryonnaire. Les oeufs non vitellins des mammifères accumulent une certaine quantité de réserves, toutefois réduites. Leur segmentation se produit lentement et le génome embryonnaire commence à être transcrit dès les premiers stades de la segmentation. Quel que soit le groupe zoologique, la transition de contrôle maternel au contrôle embryonnaire des processus développementaux ne s'effectue pas abruptement. Durant une certaine période, l'embryon utilise tant des substances codées par le génome maternel que d'autres codées par le sien, même pour une protéine donnée.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}