Laboratoire 12 : SYSTEMES FONCTIONNELS : SYSTEME SOMATOSENSORIEL
SYSTEMES MOTEURS
1- Systeme somatosensoriel
Plusieurs des structures nerveuses que vous allez étudier dans ce chapitre ont déjà été vues au cours du trimestre. Aujourd'hui, vous les étudierez dans leur contexte fonctionnel.
Les sensations somatiques, qu'elles soient extéroceptives, i.e., perçues par la peau: toucher, température et douleur, ou proprioceptives: perception de la position des parties corporelles dans l'espace et de leurs mouvements (par l'innervation des muscles, tendons et articulations), sont conscientes. Les diverses structures nerveuses responsables des sensations somatiques forment ensemble le système somatosensoriel. Le système somatosensoriel se subdivise en deux: le système lemniscal et le système antérolatéral.
Le système lemniscal véhicule la sensation du toucher de précision, si importante chez l'humain. En ce qui concerne le corps, il comprend les composantes suivantes: les terminaisons sensorielles périphériques, les nerfs périphériques, les ganglions sensoriels spinaux, les racines dorsales (afférences primaires) qui pénètrent la moelle épinière, les faisceaux gracilis et cuneatus dans les cordons dorsaux, les noyaux gracilis et cuneatus du tronc cérébral myélencéphalique, le lemnisque médian, les noyaux ventropostérieurs (VP) du thalamus, une portion de la capsule interne et, enfin, le cortex somatosensoriel. Pour la tête (la langue et certaines parties de la face surtout), il comprend: les terminaisons sensorielles périphériques, les nerfs périphériques, des ganglions sensoriels des nerfs X, IX, VII et V, le tractus trijumeau spinal (afférences primaires), les noyaux trijumeaux spinal et sensoriel principal, le lemnisque médian, les noyaux VP du thalamus, une portion de la capsule interne et le cortex somatosensoriel.
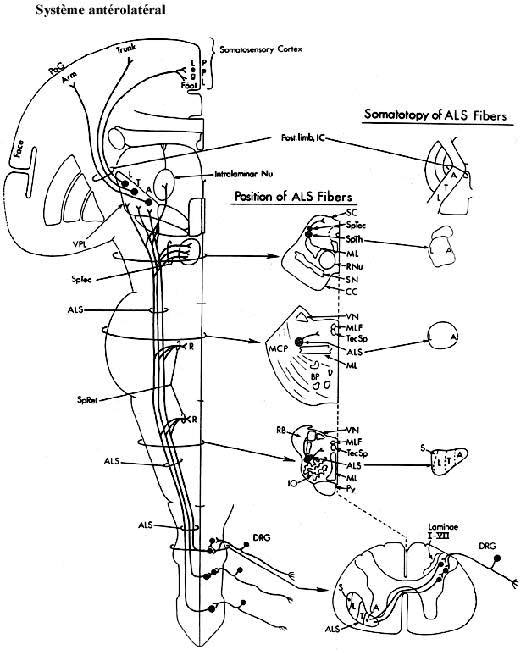
Le système antérolatéral véhicule les autres sensations extéroceptives: toucher autre que celui de précision, douleur, température, et les sensations proprioceptives: position et déplacement. Il comprend les composantes suivantes: les terminaisons sensorielles périphériques, les nerfs périphériques,Système trijumeau et du lemnisque médian les ganglions sensoriels spinaux, les racines dorsales (afférences primaires) qui pénètrent la moelle épinière, certains neurones spinaux, le faisceau antérolatéral, les noyaux VP du thalamus, une portion de la capsule interne et le cortex somatosensoriel. L'équivalent céphalique est véhiculé par le système trijumeau.
Tout au long de ces trajets, on retrouve une organisation topographique, appelée dans ce cas-ci organisation somatotopique.
- Terminaisons sensorielles:
Revisez ce qui a été dit des terminaisons sensorielles lors du premier laboratoire: histologie du système nerveux. Ensuite, allez visionner les quelques diapositives mises en démonstration qui montrent diverses terminaisons sensorielles. Passez à l'étude des lames histologiques.
La lame 7-1 montre une préparation à l'argent de la peau glabre palmaire. Repérez les terminaisons nues.
La lame 7-2 est une préparation ordinaire (H&E) du même genre de peau montrant un des types de terminaisons encapsulées: les corpuscules de Vater-Pacini qui perçoivent les vibrations. Ces corpuscules abondent dans la peau du bout des doigts chez l'humain.
La lame 7-3 montre les corpuscules de Meissner qui perçoivent le toucher de précision de la peau poilue.
- Nerf périphérique, ganglion sensoriel, racine dorsale:
Révisez ces structures que vous avez étudiées au tout premier labo et au troisième labo. N'oubliez pas que toutes les fibres du nerf, tous les neurones ganglionnaires sensoriels et tous les prolongements centraux formant la racine dorsale ne véhiculent pas les sensations du système somatosensoriel. Certains de ces éléments sont voués au système viscéral (innervation des viscères). De plus, le nerf ne contient pas que les prolongements périphériques des neurones ganglionnaires (des fibres sensorielles) mais aussi des axones moteurs.
- Moelle épinière:
Les afférences primaires pénètrent la moelle épinière par la racine dorsale. Celles véhiculant le toucher de précision se dirigent dans les cordons dorsaux et remontent la moelle jusqu'à l'encéphale. Sur le matériel de moelle épinière que vous avez utilisé durant ce trimestre (modèles, dissections, coupes histologiques), mais particulièrement sur celui de mammifères, revisez ces notions. Différenciez bien, dans le cordon dorsal, le faisceau gracilis et le faisceau cuneatus. Quelle est l'organisation somatotopique dans le cordon dorsal? Les fibres de ces faisceaux sont-elles myélinisées? Notez que la majorité des afférences cutanées qui empruntent le cordon dorsal, à leur entrée dans la moelle épinière, bifurquent et envoient une collatérale au gris spinal. Ces connexions avec la moelle épinière servent surtout dans les réflexes spinaux, et non pas au système antérolatéral (ci-après).
Les afférences primaires de la racine dorsale véhiculant les autres modalités sensorielles se dirigent dans la corne dorsale et le gris intermédiaire où elles font synapse sur des neurones spinaux. Les axones de ces neurones spinaux traversent la ligne médiane et se dirigent dans le faisceau antérolatéral, plus précisément le faisceau spinothalamique, qui remonte la moelle épinière jusqu'à l'encéphale. Qu'est-ce à dire de la somatotopie dans ce faisceau? Notez que des neurones peuvent être intercalés entre les afférences primaires et les neurones spinothalamiques.
- Noyaux des cordons dorsaux et lemnisque médian (image):
Toujours sur les préparations de mammifères, surtout les coupes histologiques, revoyez les noyaux gracilis et cuneatus, récipiendaires des faisceaux du même nom. Les neurones des noyaux gracilis et cuneatus envoient leurs axones ventralement et médialement, ces derniers traversent la ligne médiane et forment le lemnisque médian qui remonte le tronc cérébral. Dans le myélencéphale et le métencéphale, le lemnisque a une orientation dorso-ventrale, parallèle à la ligne médiane, mais notez que vers la jonction ponto-mésencéphalique le lemnisque tourne à 90o et prend une orientation médio-latérale et même oblique rostralement dans le mésencéphale. Vous pouvez particulièrement bien voir ce déplacement du lemnisque médian sur les coupes d'humain, qui sont colorées pour la myéline. Comment la somatotopie est-elle organisée tout le long du lemnisque médian?
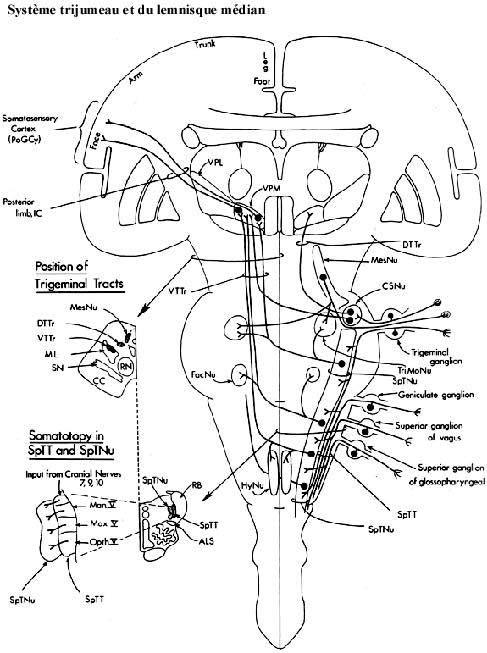
- Système trijumeau (image):
L'innervation somatique générale de la tête est fournie par les nerfs X, IX, VII et V. Nous n'avons pas de préparations qui permettent d'observer les terminaisons périphériques ou les ganglions sensoriels. Les afférences de ces différents nerfs se rassemblent en un faisceau en pénétrant le cerveau: le faisceau trijumeau spinal, qui coiffe le noyau du même nom et que vous avez observé à quelques reprises. Revoyez ce noyau sur vos préparations de mammifères et, en plus, identifiez maintenant le faisceau.
Les neurones du noyau trijumeau spinal envoient eux aussi leurs axones de l'autre côté du cerveau et ces axones vont joindre le lemnisque médian, où ils se positionnent par dessus (dorsalement) les axones provenant des noyaux gracile et cuneatus, respectant l'organisation somatotopique du lemnisque. Où se retrouveront ces fibres dans le lemnisque au niveau mésencéphalique?
- Noyaux ventropostérieurs du thalamus:
Tant le lemnisque médian que le faisceau spinothalamique aboutissent dans le groupe nucléaire médian du thalamus, plus précisément dans les noyaux appelés ventropostérieurs (médian et latéral). Les axones spinaux et lemniscaux ne convergent toutefois pas sur les mêmes neurones de VP. Encore une fois, la somatotopie est respectée dans ces noyaux. Il est possible que vos coupes histologiques de gerbille et/ou d'humain passent par ces noyaux. Comparez votre matériel avec celui des autres étudiants. Essayez aussi de localiser VP sur les coupes de cerveaux de mouton.
- Capsule interne et cortex somatosensoriel:
Les axones des neurones thalamiques empruntent la capsule interne pour gagner le cortex somatosensoriel, toujours organisés les uns par rapport aux autres de façon somatotopique.
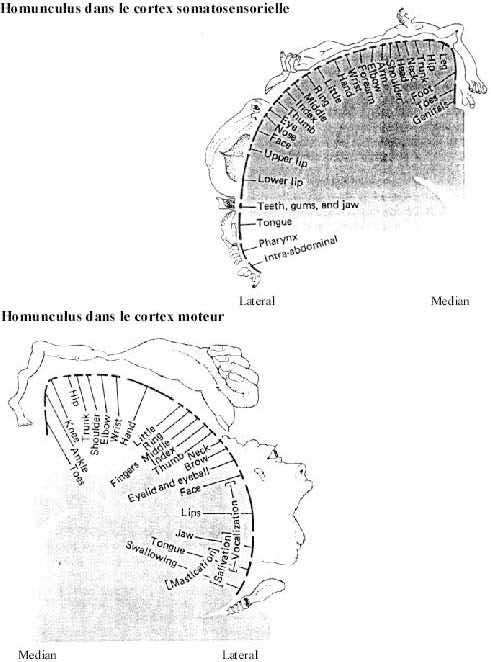
Revoyez ce qui a été étudié du cortex somatosensoriel dans les deux semaines d'étude du télencéphale. Pensez à la représentation relative des différentes parties de l'organisme (homunculus pour l'humain).
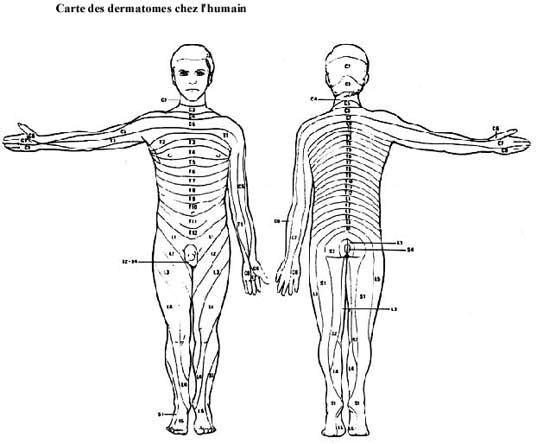
Voir schémas sur le système antérolatéral et carte des dermatomes
2- Systemes moteurs (homunculus)
Un acte moteur simple comme la locomotion, l'avancement alterné et coordonné des membres pour faire déplacer l'animal, est un mécanisme intrinsèque à la moelle épinière. Il ne requiert qu'un groupe de neurones spécifiques de la moelle: le générateur central de patron (CPG: central pattern generator, un à chaque renflement de la moelle, de chaque côté) qui commande les motoneurones innervant les membres. Il en est de même d'autres actes automatiques, comme la mastication ou la respiration qui ont leur CPG respectif dans le tronc cérébral.
Cependant, pour que l'acte moteur soit adapté à l'environnement, le CPG doit recevoir des informations sensorielles, sur la surface de marche (rugueuse ou glissante?, plane ou inclinée? etc.) ou la dureté de l'aliment, par exemple. Le CPG est donc modulé par les afférences sensorielles. De plus, l'acte moteur doit être intégré par rapport au reste de l'organisme. C'est ainsi que les CPG sont en plus modulés par différentes régions de l'encéphale qui sont appelées centres moteurs. Collectivement, on parle de système moteur.
Certains des centres moteurs encéphaliques projettent directement leurs axones à la moelle épinière (ou aux CPG du tronc):
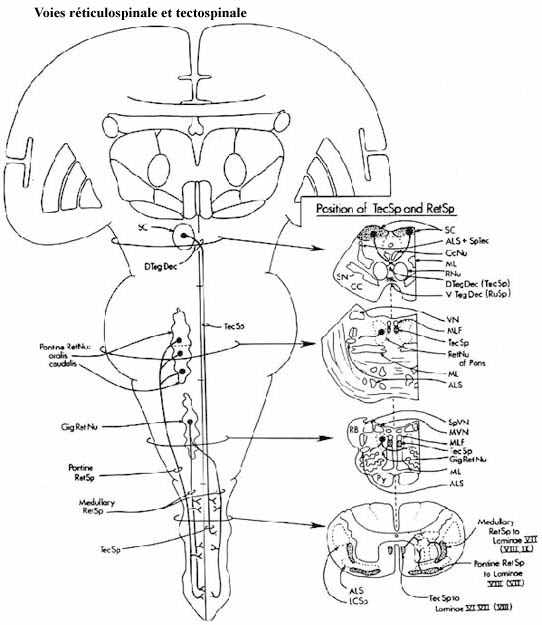
- la formation réticulée: particulièrement ses régions renfermant les gros neurones, à l'origine des voies réticulospinales. Notons aussi les sous-régions de la formation réticulée appelées raphé et locus coeruleus. Ces axones se terminent dans toutes les régions de la matière grise spinale. Les connexions réticulospinales existent chez tous les vertébrés.
Le rôle de la formation réticulée dans le contrôle de la motricité est mal compris. Recevant des inputs de nombreuses sources et contenant des neurones utilisant toute une panoplie de neurotransmetteurs, la formation réticulée est un centre intégrateur. Elle joue un rôle dans les cycles éveil-sommeil et est notamment impliquée dans la phase du sommeil appelée sommeil paradoxal qui correspond aux épisodes de rêves (même si vous rêvez que vous courrez, vous restez néanmoins immobile dans votre lit parce que des neurones réticulés sérotoninergiques inhibent directement les motoneurones spinaux innervant les membres). Son rôle est excitateur dans certains cas, inhibiteur dans d'autres. Elle influence directement la locomotion et les autres fonctions motrices.
- le complexe vestibulaire: les noyaux latéral, médian et inférieur, à l'origine des voies vestibulospinales qui se terminent principalement dans la région médiane de la corne ventrale. Les connexions vestibulospinales existent chez tous les vertébrés.
Les noyaux vestibulaires reçoivent les afférences primaires de la portion vestibulaire du nerf VIII qui innerve les organes de l'équilibre dans l'oreille interne. Ils influencent donc les mouvements du cou et des membres en réponse à l'accélération linéaire (gravité) et angulaire (mouvements de la tête dans l'espace) et contribuent au maintien de la posture et de l'équilibre au repos comme durant la marche.
Les noyaux vestibulaires, notamment le noyau supérieur, se projettent aussi aux noyaux oculomoteurs et contrôlent le mouvement des yeux.
- le noyau rouge: à l'origine de la voie rubrobulbaire et rubrospinale qui se termine principalement dans la formation réticulée entourant les noyaux moteurs du tronc ainsi que dans la région médiane de la corne dorsale. Les connexions rubrospinales se retrouvent au moins à partir des poissons téléostéens.
Le rôle exact de ce noyau dans les comportements moteurs est obscur. Il est interconnecté à plusieurs autres centres du système moteur.
- le tectum: ses gros neurones profonds, à l'origine de la voie tectospinale, envoient leurs axones principalement à la région médiane de la corne ventrale. La voie tectospinale est aussi ancienne phylogénétiquement.
Le tectum est fortement impliqué dans la coordination des mouvements des yeux et de la tête mais en réponse à des stimuli visuels, comparativement au complexe vestibulaire qui accomplit une tâche similaire en réponse aux stimuli de l'oreille interne.
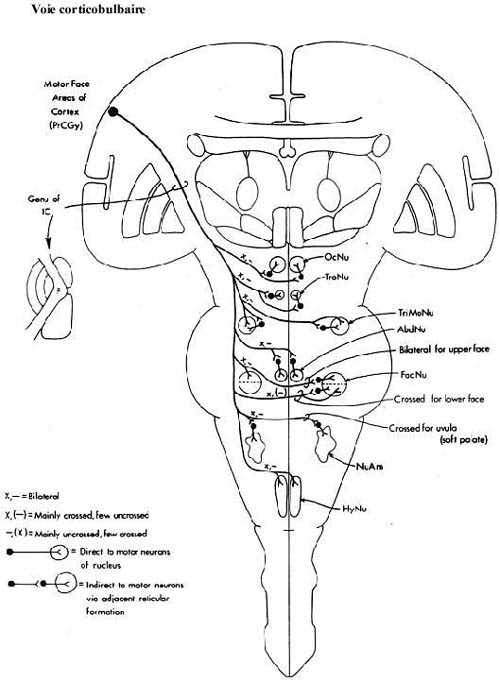
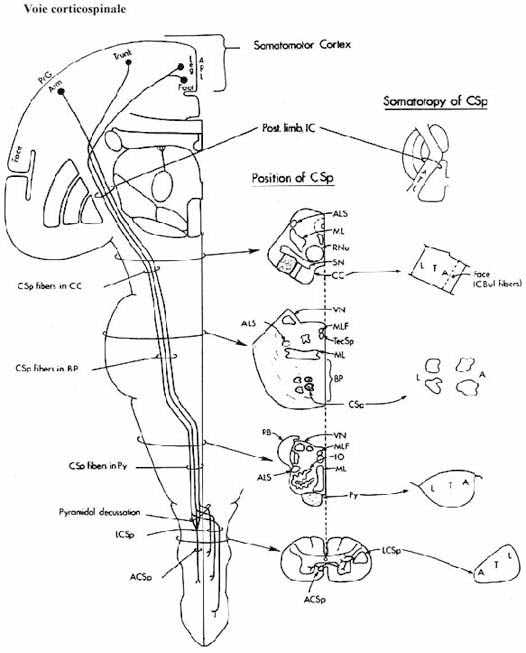
- le cortex cérébral: les aires motrice, prémotrice et somatosensorielle, à l'origine de la voie corticobulbaire et corticospinale (pyramidale) acheminent leurs axones aux alentours des noyaux moteurs du tronc et dans la corne dorsale de la moelle, surtout latéralement. La voie corticospinale n'est présente que chez les mammifères. Même chez les marsupiaux, elle est réduite. Il y a chevauchement entre les champs de distribution des axones rubraux et corticaux. Les projections directes du cortex aux motoneurones (corne ventrale) se font plus nombreuses en montant l'échelle phylogénétique mammalienne. Le cortex se projette aussi au noyau rouge et aux noyaux du pons (voir ci-bas).
On dit que le cortex cérébral contrôle les aspects volontaires des mouvements. Son ablation résulte en spasticité (rigidité) et en parésie (paralysie incomplète). Comme le cortex projette ses axones aux centres moteurs précédents qui sont à l'origine d'importantes projections à la moelle épinière, la perte de fonction corticale pourra aussi affecter tous ces systèmes descendants.
Revoyez ces centres, la localisation de leurs axones dans la moelle épinière et leur site de terminaison respectif dans la moelle. Rappelez-vous que ces systèmes de projection, particulièrement celui provenant du cortex, sont organisés somatotopiquement.
D'autres centres moteurs encéphaliques ne sont pas connectés directement aux centres moteurs primaires. Ils sont plutôt reliés aux centres moteurs encéphaliques dont nous venons de parler, de même qu'au thalamus et entre eux.
- le cervelet: il reçoit beaucoup d'information proprioceptive de la moelle épinière et du tronc cérébral lui communiquant l'état d'étirement des muscles et tendons, ceci par les voies spinocérébelleuses.
De plus, le cervelet reçoit des afférences de noyaux du tronc appelés noyaux précérébellaires: le noyau cuneatus latéral (ou externe) et le noyau réticulé latéral, que vous n'avez pas étudiés précédemment, et l'olive inférieure que vous avez déjà vue. Ces relais précérébellaires reçoivent eux aussi de l'information proprioceptive de la moelle et du tronc.
Les noyaux du pons, que vous avez étudiés, sont eux aussi précérébellaires mais ne reçoivent pas de la moelle épinière. Leurs afférences leur proviennent de toutes les aires du cortex cérébral, et pas seulement des aires qui innervent la moelle (i.e., motrice, prémotrice et somatosensorielle).
Revoyez toutes ces structures que vous avez déjà étudiées ainsi que, si présents sur vos coupes, les noyaux cuneatus latéral et réticulé latéral. Revoyez le chapitre sur le cervelet et demandez-vous quel pédoncule cérébelleux empruntent toutes ces différentes afférences au cervelet, à quel type de fibres appartiennent-elles: moussues ou grimpantes, où se terminent-elles dans le cervelet: noyaux profonds, cortex (quelle couche?).
Les cellules de Purkinje du cortex cérébelleux envoient leurs axones vers les noyaux profonds. Par les pédoncules cérébelleux supérieurs, les noyaux intermédiaire (interposé) et latéral (dentelé) atteignent deux cibles principales: le noyau rouge et le groupe nucléaire latéral du thalamus en plus d'un gros noyau intralaminaire (le centromédian). Le noyau cérébellaire médian (fastigial) innerve les segments cervicaux de la moelle épinière pour influencer les mouvements du cou (donc de la tête) en plus du complexe vestibulaire. Ce dernier reçoit également une projection cérébellocorticale directe, depuis de lobe noduloflocculaire.
Les fonctions du cervelet dans le contrôle moteur sont nombreuses et encore mal comprises de nos jours, malgré l'abondance d'études morphologiques, physiologiques et pharmacologiques effectuées sur cet organe. Les lésions cérébelleuses résultent en ataxie (mouvements non coordonnés) et en tremblement. On dit que le cervelet est important dans le contrôle fin des mouvements et dans l'apprentissage moteur. Par exemple, vous avez besoin de votre cervelet pour jouer du piano.
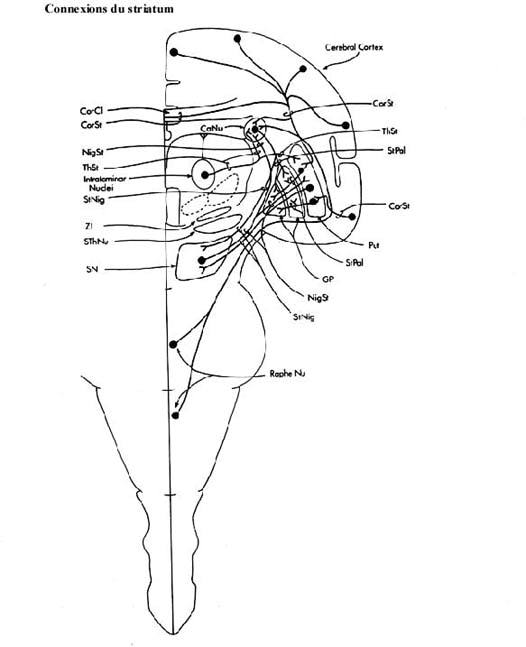
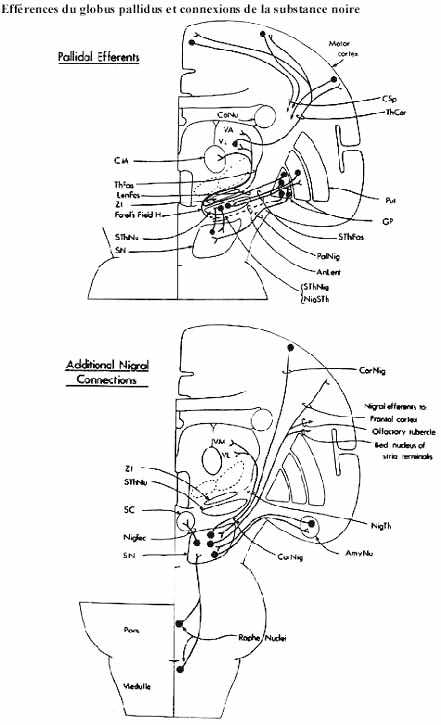
- les noyaux basaux: le caudé et le putamen, formant ensemble le striatum, reçoivent massivement de toutes les aires du cortex cérébral (non seulement celles projetant à la moelle épinière), selon une organisation topographique. Ils projettent eux-mêmes leurs axones au globus pallidus, surtout, et à la substance noire. La connexion avec la substance noire est réciproque. Les axones nigrostriataux sont dopaminergiques. Leur destruction est la cause de la maladie de Parkinson, caractérisée par un tremblement régulier. Au contraire, une trop grande activation de la voie nigrostriatale serait à l'origine de la maladie de Huntington (danse de Saint-Guy), caractérisée par l'apparition de mouvements choréiformes.
- le globus pallidus, que vous avez vu dans le cadre du sous-thalamus mais, tel qu'expliqué, que certains auteurs incluent dans les noyaux basaux, constituent la porte de sortie du striatum. Son origine est néanmoins diencéphalique. Les axones pallidaux sont dirigés vers le thalamus, plus précisément vers les noyaux du groupe latéral et le noyau intralaminaire centromédian récipiendaires d'afférences cérébelleuses. L'un de ces noyaux thalamiques projette ses axones aux aires motrice et prémotrice du cortex cérébral. Le noyau centromédian envoie ses axones au striatum.
Le globus pallidus envoie aussi des axones au noyau sous-thalamique. En fait cette connexion est réciproque. La destruction du noyau sous-thalamique amène l'expression de mouvements balistiques des membres contralatéraux au noyau lésé.
Dans l'ensemble, les déficits des noyaux basaux appartiennent à la catégorie des dyskinésies: expression de mouvements involontaires, sans raison, mais pas de paralysie.
Revisez les labos des semaines précédentes. L'examen final de T.P. aura lieu la semaine prochaine. Il comprend 40 questions d'identification et n'est pas cumulatif. Toutefois, certaines des notions apprises avant l'intra ont été revues dans un contexte différent. Elles sont sujettes à examen. Fiez-vous au cahier pour savoir ce qui doit être appris depuis l'intra.
BONNE ETUDE ET BONNE CHANCE A L'EXAMEN
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}