 Jean-Pierre Raynauld, Biophysicien
Jean-Pierre Raynauld, Biophysicien
Université de Montréal
One can
browse this page in english
Se puede visionar esta pagina en castellano
Pode-se também visualizar esta página em português
Autres pages qui peuvent vous intéresser:
A: Un convertisseur d'unités de lumière Light convert
Voici la page Web d'un chapitre qui vient de paraître dans les " Proceedings of the NATO ASI on Neurobiology", qui a eu lieu à Erice, en Sicile, du 2 au 12 mai, 1995, " Neurobiology: Ionic Channels, Neurons, and the Brain: édités par V. Torre et F. Ponti", Plenum Press,1996. Ceux qui sont intéressés à plus de détails peuvent en faire la demande du chapitre " A Compartment Model for Vertebrate Phototransduction Predicts Sensitivity and Adaptation" pp. 201-215 en m'écrivant à l'adresse apparaissant au bas de cette page. Je leur ferai parvenir un tiré-à-part. Les versions Postcript et PDF sont disponibles électroniquement. Version Postcript ou Version PDF
Netscape 2.0 est le logiciel de choix pour voir ce document. La sélection des icônes fait apparaître des images ou des graphiques élargis.
Un modèle à compartiments de la phototransduction chez les vertébrés
Résumé
Les propriétés différentielles des cônes et des bâtonnets en regard de la sensibilité et de l'adaptation sont expliquées par un modèle à compartiments. Pour les cônes, le compartiment naturel est le pli de la membrane plasmatique; pour les bâtonnets le compartiment a été défini de façon empirique. Dans ce modèle, le nombre de compartiments contrôle l'amplitude de la réponse à l'action d'un photon; la taille du compartiment détermine la position de la courbe intensité-réponse sur l'axe des intensités. En utilisant le théorème d'ergodicité , il a été possible de prédire les propriétés en adaptation à partir du nombre de compartiments et de la constante de temps de la phase décroissante de la réponse à un photon.
Introduction
 Je
suis un biophysicien dont les intérêts sont présentement de trouver pourquoi
les cônes et les bâtonnets des vertébrés possèdent des sensibilités et des
propriétés d'adaptation différentes, alors que la biochimie, qui sous-tend ces
phénomènes, semble très similaire dans ces deux classes de photorécepteurs [1].
Je
suis un biophysicien dont les intérêts sont présentement de trouver pourquoi
les cônes et les bâtonnets des vertébrés possèdent des sensibilités et des
propriétés d'adaptation différentes, alors que la biochimie, qui sous-tend ces
phénomènes, semble très similaire dans ces deux classes de photorécepteurs [1].
Comme point de départ, j'utilise les différences anatomiques connues dans la structure du segment externe des photorécepteurs magnifiquement illustrées par Alfieri, R. et al. [2] (permission demandée).
Figure 1: Bâtonnet montrant
l'empilement des disques dans une coquille cylindrique.
 Au niveau du segment externe , la différence principale
entre les cônes et les bâtonnets réside dans le fait que les disques de cônes
sont des replis de la membrane plasmatique, alors que, chez les bâtonnets, les
disques sont fermés et empilés dans une coquille cylindrique.
Au niveau du segment externe , la différence principale
entre les cônes et les bâtonnets réside dans le fait que les disques de cônes
sont des replis de la membrane plasmatique, alors que, chez les bâtonnets, les
disques sont fermés et empilés dans une coquille cylindrique.
De plus, une étude tridimensionnelle [3] montre que les cônes sont des structures presque entièrement fermées sur elles-mêmes avec une ouverture d'environ 20 degrés sur la circonférence.
Figure 2: Illustration de cone
montrant que les plis sont partie intégrante de la membrane plasmatique.
Ce modèle propose l'existence de compartiments qui limitent en volume l'étendu des réactions biochimiques conséquentes à la capture d'un photon. Chez le cône, je veux démontrer que les membranes formant le pli déterminent également la grandeur du compartiment. Chez le bâtonnet, nous faisons face à un compartiment dit "opérationnel", que je définirai plus loin. Deux facteurs et un hypothèse sont importants pour la discussion qui va suivre:
- Le temps de montée de la
réponse à un photon.
- Le coéfficient de
diffusion des molécules impliquées dans la cascade biochimique.
- Dans un compartiment donné, l'action d'un seul photon est maximale.
Intensité-Réponse
 La biochimie
responsable de la phototransduction a été l'objet d'études approfondies au
cours des quinze dernières années. Le résultat est bien illustré dans ce schéma
de Nakatani.. L'idée que le segment externe des
photorécepteurs est compartimenté n'est pas nouvelle. Dès 1981, dans le but
d'expliquer la forme de la courbe des données "Intensité-Réponse"
Lamb et al. [4] ont
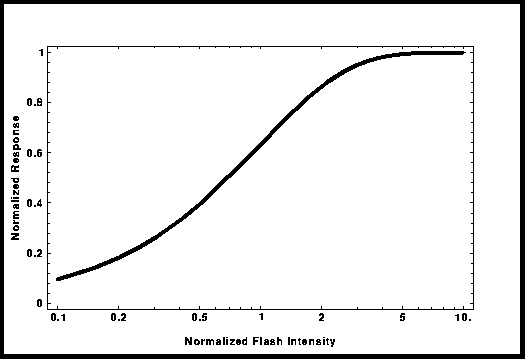
developpé le modèle de "l'occlusion totale". La courbe résultante
dite de "Saturation exponentielle" peut être utilisée pour remplacer
la courbe de "Michaelis-Menten"; son expression mathématique est :
R/Rmax = 1 - Exp[-kI], où I est l'intensité du flash et k, un paramètre
d'ajustement. Dans ce modèle, je vais tenter d'obtenir le nombre de
compartiments et leur taille. Des résultats, j'obtiendrai la réponse à un
photon et la valeur du paramètre "k".
La biochimie
responsable de la phototransduction a été l'objet d'études approfondies au
cours des quinze dernières années. Le résultat est bien illustré dans ce schéma
de Nakatani.. L'idée que le segment externe des
photorécepteurs est compartimenté n'est pas nouvelle. Dès 1981, dans le but
d'expliquer la forme de la courbe des données "Intensité-Réponse"
Lamb et al. [4] ont
developpé le modèle de "l'occlusion totale". La courbe résultante
dite de "Saturation exponentielle" peut être utilisée pour remplacer
la courbe de "Michaelis-Menten"; son expression mathématique est :
R/Rmax = 1 - Exp[-kI], où I est l'intensité du flash et k, un paramètre
d'ajustement. Dans ce modèle, je vais tenter d'obtenir le nombre de
compartiments et leur taille. Des résultats, j'obtiendrai la réponse à un
photon et la valeur du paramètre "k".
Figure 3: Courbe
"Intensité-Réponse
dans le modèle "Occlusion totale".
Adaptation Weber-Fechner
Sous une
illumination dont l'intensité n'est pas suffisante pour réduire la densité du
pigment visuel, la réduction de sensibilité suit la loi dite de
"Weber-Fechner", énoncée il y plus d'un siècle. Cette loi montre que
la sensibilité normalisée décroît avec l'intensité de la lumière ambiante
"I" selon la formule toute simple " Io/(I + Io) ", où Io
est la lumière ambiante requise pour réduire la sensibilité par un facteur
d'une demie.
Il est cependant
intéressant de noter qu'un siècle avant Weber et Fechner, Pierre Bouguer avait
été le premier à mesurer l'effet du la lumière ambiante sur le seuil de
détection en milieu photopique. Il avait montré qu'une variation de la lumière
d'environ 1/64 était requise pour engendrer la détection.
Les propriétés d'adaptation
d'un modèle à compartiments n'ont pas fait l'objet d'études antérieures. Mon
analyse est du type "boite noire"; après une isomérisation qui
perturbe le compartiment, les réactions contrôlant la réponse retournent à la
situation d'équilibre qui existait à l'obscurité. Parmi toutes les réactions
chimiques qui participent à ce processus, une d'entre elles est limitante et je
fais l'hypothèse que le forme du photo-courant reflète cette réaction qui
contrôle la sensibilité. Il importe donc de définir la forme de la réponse à un
photon et j'ai choisi l'exponentielle. Dans ce schéma, suite à une
isomérisation, le courant subit une montée rapide suivie par une décroissance
exponentielle, caractérisée par une constante de temps. Ceci n'est pas une
mauvaise approximation car plusieurs cônes et bâtonnets ont de telles réponses [5].
Dans mon analyse d'un
système à compartiments, je fais appel à un théorème de mécanique statistique
nommé " théorème d'ergodicité" qui s'énonce ainsi: Dans un
procédé stochastique stationnaire, un grand nombre d'observations faites sur un
seul système à N moments arbitraires possèdent les mêmes propriétés
statistiques que les observations faites sur N systèmes à un moment donné.
S'il m'est permis de faire une équivalence entre compartiment et système, je
peux faire l'analyse d'un compartiment sous une lumière donnée et prédire le
comportement du photorécepteur qui, dans son entier, est une structure formée
d'un certain nombre de compartiments.
Suite à une isomérisation
qui produit une réponse maximale ( normalisée ici à 1) et décroissant
exponentiellement selon une constante de temps "T", l'amplitude de la
réponse à un second photon dépend du temps depuis le photon précédent et sera
égal à " 1 - Exp[-t/T] ". Étant donné que les intervalles entre les
isomérisations obéissent à la loi de Poisson, il est possible de simuler une
intensité ambiante donnée et de trouver la réponse moyenne du compartiment.
Ceci fut fait à l'aide de Mathematica et les résultats apparaissent
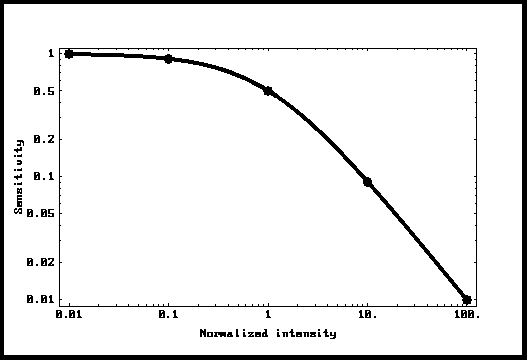
comme les points de la Fig. 4. On peut voir que les points se superposent
exactement sur la courbe Io/(I+Io). La conclusion est donc qu'un tel système se
comporte exactement comme le prévoit la loi Weber-Fechner. Ceci représente
une contribution majeure de cette analyse
Figure 4: Sensibilité en fonction de la
lumière ambiante
La courbe Weber-Fechner .
Le paramètre Io, qui à
date, servait uniquement à ajuster la courbe aux données expérimentales,
possède maintenant une signification toute particulière liée à la structure du
segment externe, c'est à dire au nombre de compartiments et à la constante de
temps de la réponse à un photon. La relation étant que Io est égal à N/T, où N
est le nombre de compartiments et T, la constante de temps de la décroissance
exponentielle.
Le résultat final est donc
que, de la connaissance de la grandeur du compartiment, du nombre de
compartiments, et de la constante de temps associée à la réponse à un photon,
on peut prédire l'amplitude de la réponse unitaire, et localiser de façon
absolue la position des courbes "Intensité-Réponse" et
"Weber-Fechner" sur leur axe "Intensité" respectif.
Résultats et Discussion
Les cônes
D'une part,
la réponse des cônes de mammifères à un flash de faible intensité pointe avec
une latence d'environ 50 msc.; d'autre part, l'iodopsine, la transducine et la
phosphodiesterase, protéines membranaires impliquées dans la réaction en
cascade, ont des coefficient de diffusion entre 0.5 et 2 microns carrés par
sec.. Diffusant dans un espace à deux dimensions, très peu de ces molécules
activées peuvent quitter le pli membranaire dont le diamètre est de 1.5 à 2
microns dans un si court lapse de temps. Pour le cGMP, qui diffuse en solution
et dont le coefficient de diffusion est plus grand, il faut considérer dans
quelle mesure, des molécules situées dans des disques adjacents au disque où
l'isomérisation s'est produite quittent leur disque pour venir se faire
hydrolyser dans le disque actif. Je fais l'hypothèse que, faute de temps, cela
est un événement rare. A l'intérieur du disque actif, on peut faire un calcul
simple et montrer que le nombre de phosphodiestérases activées peut excéder le
nombre de molécules libres de cGMP. Il existe donc une possibilité réelle
qu'une seule isomérisation cause la fermeture de tous les canaux ioniques d'un
disque ou d'un pli de cône.
Le premier résultat de
cette analyse est que, si dans les cônes, le disque forme un compartiment et
que la réponse à l'intérieur de ce compartiment est maximale, alors l'amplitude
de la réponse à un photon est égale à 1/N, où N est le nombre de disques. La
microscopie électronique nous dit que le nombre de disques par micron est
d'environ 33. Le nombre total de disques est donc égal à la longueur du segment
externe multiplié par 33. Dans le cas du cône de tortue, ce nombre est environ
500; la réponse prédite suite à l'action d'un photon est un changement de 0.2%,
la valeur trouvée expérimentalement est de 0.16% [6] en accord avec la prédiction. Des données
obtenues en 2006 [7] offrent un
support additionnel au model 1/N. Chez des souris modifiées
génétiquement on peut obtenir des cones avec des segments externes de longueur variable
Chez les souris Gt alpha -/- les cones S et M ont des segments externes d'une longueur
de 13.4 microns et ont respectivement une réponse
à un photon de 0.2 et 0.22 % de la réponse maximale.
Le model 1/N prédit 1/(13.4 x 33) ou 0.23 %. Chez
les souris Nrl -/- les cones S ont un segment externe de 7.1 microns et la réponse à un photon est 0.44%,
le model prédit 1/(7.1 x 33) ou 0.43 %. il est difficile de demander un meilleur
accord entre des données expérimentales
et un modèle théorique. Un autre
résultat issu de cette analyse est une hypothèse pour expliquer pourquoi les
cônes sont généralement courts. En effet, si l'ajout d'un disque augmente la
probabilité de capture d'un photon, la réponse, elle, diminue de 1/N à 1/(N+1).
La nature a donc du faire un compromis entre " capturer des photons"
et "donner une réponse adéquate à la capture de ce photon".
Connaissant les dimensions
du compartiment, c'est-à-dire du disque ou pli plasmatique, il est possible de
calculer la valeur du paramètre "k" dans la formule de saturation
exponentielle [8]. De
la valeur de la densité optique du segment externe et de celle de l'efficacité
quantique, on trouve que "k" est égal à: 0.00059 fois le carré du
diamètre du segment externe exprimé en microns. Lors de la dérivation de cette
formule, on a considéré que le cône était stimulé de façon transverse, ce qui
correspond à la situation expérimentale. Cette dérivation est également valide
pour une stimulation axiale correspondant à la situation physiologique en
autant que le segment externe n'est pas trop long. Dans le cas des cônes de Macaca
le diamètre est d'environ 1.5 micron, la valeur de "k" calculée donne
0.001 micron carré, alors que la valeur obtenue expérimentalement est de 0.0004
[9], soit environ la
moitié.
Un estimé de l'intensité
(mesurée en isomérisations par sec.) nécessaire pour réduire la sensibilité de
50 % est obtenu en divisant le nombre de compartiments (800 pour le cône de
tortue) par la constante de temps qui caractérise la décroissance de la réponse
à un flash de faible intensité. Ce calcul donne environ 7300 pour la valeur de
Io. Expérimentalement, on trouve des valeurs entre 2000 et 16000 par sec. [10].
Les bâtonnets
Dans le cas
des bâtonnets, le compartiment n'est pas défini par des membranes comme ce fut
le cas pour les cônes mais par la combinaison du fait que les protéines
activées dans la phototranduction ont toutes un temps de vie limité et que le
CGMP qui contrôle l'ouverture des canaux possède un coefficient de diffusion
réduit dans l'environnement du segment externe. La réponse d'un bâtonnet de
mammifère pointe à environ 100 msec., ceci permet l'activation de plus de
molécules de transducine et de phosphodiestérase. L'action de la biochimie peut
quitter l'espace inter disque et s'étendre longitudinalement. La géométrie de
l'empilement des disques dans le segment externe des bâtonnets est importante. Pour
se propager longitudinalement, la réduction de cGMP doit réduire la
concentration entre chaque disque le long de son parcours. Ceci limite l'action
de la biochimie. Nous sommes en présence d'un compartiment dit
"opérationnel".
Des calculs complexes en dynamique
des fluides seraient requis pour déterminer la concentration longitudinale de
cGMP dans le voisinage de la membrane plasmatique suite à une isomérisation
dans un disque. Toute fois, une formule empirique a été obtenue de l'analyse de
la réponse à un photon chez plusieurs espèces. Cette relation dit que la
surface de membrane plasmatique où les canaux ioniques sont fermés est égale à
celle d'un disque (deux feuillets) du segment externe. Un simple calcul donne
que la longueur impliqué doit être égale au diamètre divisé par deux. Pour les
bâtonnets de grenouille, de rats, de singes et aussi humains, la réduction de
courant prédit est de 4-5%, soit la valeur trouvée expérimentalement.
Des calculs similaires à
ceux faits pour les cônes donnent une valeur de "k" pour les
bâtonnets égal à 0.01 fois le cube du diamètre du segment externe exprimé en
microns. Ce calcul permet de localiser la courbe "Intensité-Réponse"
de façon absolue sur l'axe de l'intensité. Pour les bâtonnets des mammifères,
la valeur calculée est de 0.08 microns carrés; les valeurs obtenues
expérimentalement varient entre 0.006 et 0.07 [8,11].
Le nombre de compartiments
"N" d'un bâtonnet est obtenu en divisant la longueur totale du
segment externe par d/2, ceci donne un nombre près de 20. Io dans la relation
de Weber-Fechner devient alors 20 divisé par la constante de temps de la
décroissance exponentielle. Pour les bâtonnets de vertébrés supérieurs (homme,
singe), la prédiction donne un Io entre 110-113 isomérisations par sec. alors
que les résultats expérimentaux donnent entre 100 et 120 par sec. [8,11]
Conclusion
L'analyse
de la structure géométrique du segment externe des photorécepteurs des vertébrés
a donné des résultats intéressants.
- Dans un premier temps, il a été possible
de prédire la réponse à un photon. et de calculer la valeur du paramètre
"k" dans la formule de "saturation exponentielle",
permettant ainsi de localiser la courbe sur l'axe des intensités.
- En second, à l'aide du théorème
d'ergodicité, il a été possible de prédire les propriétés d'adaptation
d'un système à compartiments. Si la réponse est maximale et la
décroissance exponentielle, le résultat est la loi de Weber-Fechner. Cette
courbe est aussi localisée de façon absolue sur l'axe des intensités.
De plus la validité du
modèle peut être testée. En effet, chez les vertébrés à sang froid, la
cinétique de la réponse à un flash varie avec la température[12]. On peut donc voir si les propriétés
d'adaptation (Io) changent selon le modèle présenté ci-haut.
Vos commentaires sont
appréciés-courrier électronique: jean-pierre.raynauld@umontreal.ca
1. Lamb
TD, Pugh EN (1992) G-protein cascades: gain and kinetics . Trends Neurosci 15:
291-298.
2. Alfieri
R, Sole P, Gentou C, Kantelip B, Kantelip J-P . (1984) Les cellules
rétiniennes. Ed. D.G.P.L. Paris.
3.
Eckmiller MS (1987) Cone Outer Segment Morphogenesis: Taper Change and Distal
Invaginations. J Cell Biol 105: 2267-2277.
4. Lamb
TD, McNaughton PA, Yau KW (1981) Spatial spread of activation and background
desensitization in toad rod outer segments. J Physiol 319: 463-496.
5. Corson
DW, Cornwall MC, MacNichol EF, Tsang S, Derguini F, Crouch RK,Nakanishi K
(1994) Relief of opsin desensitization and prolonged excitation of rod
photoreceptors by 9-desmethylretinal. Proc Natl Acad Sci USA 91: 6958-6962.
6. Baylor
DA, Hodgkin AL (1973) Detection and resolution of visual stimuli by turtle
photoreceptors. J Physiol 234: 163-198
7.
Nikonov SS, Kholodenko R, Lem J, and Pugh EN (2006)
Physiological features of the S- and M-cone Photoreceptors of wild-type mice
from single-cell recordings. J.Gen. Physiol. 127(4), 359-374
8. Baylor
DA., Nunn BJ, Schnapf JL (1984) The photocurrent noise and spectral
sensitivities of rods of the monkey Macaca fascicularis. J Physiol 357: 575-607
9. Schnapf
JL, Nunn BJ, Meister M, Baylor DA (1990) Visual transduction in cones of the
monkey Macaca fascicularis. J Physiol 427: 681-713
10. Baylor
DA, Hodgkin AL (1974) Change in time scale and sensitivity in turtle
photoreceptors. J Physiol 242: 729-758
Burkhardt DA (1994) Ligth adaptation and photopigment bleaching in cone
photoreceptors in situ in the retina of the turtle. J Neurosci 14(3): 1091-1105
11. Kraft
TW, Schneeweis DM, Schnapf JL (1993) Visual Transduction in human
photoreceptors. J Physiol 464: 747-765
12. Lamb
TD (1984) Effects of temperature changes on toad rod photo currents. J Phsysiol
346: 557-578
Issue de mon bocal à poissons rouges
Jean-Pierre Raynauld
(dernière mise à jour, le 24 mai 2007)