 Jean-Pierre Raynauld, Biofísico
Jean-Pierre Raynauld, Biofísico
Universidade de Montreal
You can
also visit my page in english
On peut aussi visionner cette page en français
Se puede visionar esta pagina en castellano
Esta é uma versão em Web Page de um modelo que faz parte dos Proceedings of the NATO ASI on Neurobiology , ocorrido em Erice, Sicília, de 2 a 12 de maio de 1995, e que foi publicado pela Plenum Press: "Neurobiology: Ionic Channels, Neurons, and the Brain ,V. Torre and F. Ponti, eds, 1996". Os interessados em mais detalhes podem pedir uma cópia do capítulo "A Compartment Model for Vertebrate Phototransduction Predicts Sensitivity and Adaptation, pp. 201-215" ou fazer um download da versão em Postcript ou da versão em PDF do manuscrito completo (seu browser deve estar apropriadamente configurado para esta operação). Os pedidos de manuscritos serão respondidos por e-mail.
Este documento é melhor visualizado utilizando-se Netscape 2.0 ou outro mais recente. Clicando nas figuras pequenas ou nos números sublinhados pode-se visualizar uma imagem colorida maior ou um gráfico em tamanho maior.
Um modelo compartimentalizado da fototransdução nos vertebrados
Resumo
Desenvolvemos um modelo compartimentalizado para explicar as diferentes propriedades de adaptação e sensibilidades dos cones e bastonetes dos vertebrados. Para os cones, o compartimento natural é uma câmara simples com a membrana plasmática; para os bastonetes, o tamanho do compartimento deve ser definido empiricamente. Neste modelo, o número de compartimentos controla a amplitude da resposta de cada fóton; o tamanho do compartimento determina a posição da curva de Resposta/Intensidade no eixo de intensidade. Usando o teorema da ergodicidade (ou teorema ergódico), as propriedades de adaptação podem ser previstas a partir do número de compartimentos e da constante de tempo de queda da resposta de cada fóton.
Introdução

Sou biofísico, e atualmente estou interessado em saber por quê bastonetes e cones na retina dos vertebrados têm diferentes sensibilidades à luz e por quê as suas propriedades de adaptação sob condições normais de luz são também tão diferentes quando a bioquímica, bem revisada em [1], parece muito semelhante nestes dois tipos de receptores.
Como ponto de partida, utilizo a já bem estudada diferença anatômica entre a estrutura do segmento externo destes fotoreceptores, e para ilustrar isto utilizo as ilustrações de Alfieri, R. et al. [2] (permissão obtida).
Figura 1: Bastonete mostrando o
empilhamento dos discos numa concha cilíndrica.

A principal diferença é que, enquanto os discos dos cones ficam livres no espaço, os discos dos bastonetes são fechados dentro de uma concha cilíndrica.
Mais ainda, um estudo tridimensional mostra que o disco de um cone é conectada à parte ciliar da ultraestrutura através de uma abertura mínima que tem menos do que 20 graus do círculo [3].
Figura 2: Figura de um cone mostrando
os discos como sendo parte da membrana plasmática.
A próxima consideração envolve a existência de compartimentos nestas estruturas que limitam, em volume, a extensão das reações bioquímicas que se seguem à captura de um fóton. No caso de um cone, tentarei mostrar que as memebranas limitam o tamanho do compatimento; quando, para o bastonete, estamos lidando com um "compartimento operacional", que definirei mais tarde. Dois fatores e uma hipótese são importantes para a discussão a seguir:
- Tempo para o pico da
resposta a um único fóton.
- Coeficiente de difusão
das moléculas envolvidas na cascata bioquímica.
- A ação de um fóton é
máxima num dado compartimento.
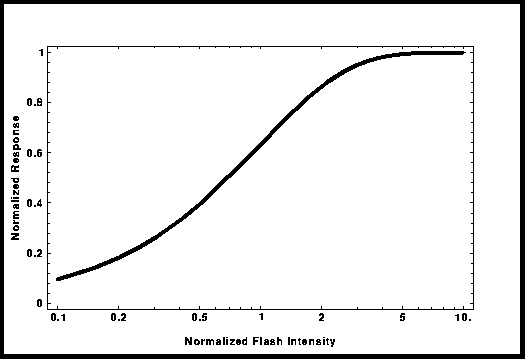
Intensidade-Resposta
A bioquímica que está sob a fototransdução tem sido foco de estudos nos
últimos quinze anos; o que se sabe atualmente é bem ilustrado por um esquema de
Nakatani. A idéia de que o segmento externo é
compartimentalizado não é nova. Já em 1981, para explicar a forma dos dados na
curva intensidade/resposta, Lamb et al. [4] desenvolveram o modelo de oclusão total. Esta curva,
também chamada de "saturação exponencial", pode ser utilizada para
modelar dados experimentais e substituir a curva "Michaelis-Menten";
a sua expressão matemática é: R/Rmax = 1 - Exp[-kI], onde I é a intensidade do
flash e k o parâmetro de acerto na curva. Neste modelo, tento obter uma
estimativa para o número de compartimentos e seu tamanho tanto em bastonetes
quanto cones. A partir dos resultados, tenho uma estimativa da resposta para
cada fóton individualmente e um valor para "k" .
Figure 3: Forma da curva de
intensidade-resposta
no "modelo de oclusão total".
Adaptação Weber-Fechner
Sob uma iluminação cuja
intensidade não é suficiente para reduzir a densidade do pigmento visual, a
redução de sensibilidade dos fotoreceptores com o aumento da luz ambiente segue
uma lei descrita há mais de um século, chamada de lei de Weber-Fechner. Ela
mostra que a sensibilidade normalizada cai com o aumento da intensidade da luz
ambiente "I" de acordo com a relação simples " Io/(I + Io)
", Io sendo a intensidade necessária para reduzir a sensibilidade pela
metade.
É interessante notar que um
século antes de Weber e Fechner, o francês Pierre Bouguer foi o primeiro a
medir o efeito da luz ambiente no limiar do estado fotópico. A medida que ele
fez do limiar incremental foi de 1/64 com relação à intensidade de fundo, um
valor ainda aceito hoje.
As propriedades de adaptação
de um modelo compartimentalizado nunca foram analisadas anteriormente. Meu
enfoque é o assim chamado "caixa preta" (black box), e segue as
seguintes diretrizes gerais: Após uma isomerização que produziu um efeito
máximo, o compartimento é perturbado e as reações que controlam a resposta
estão sendo restauradas para os valores de equilíbrio de quando o compartimento
está adaptado ao escuro. Das muitas reações que ocorrem através deste processo,
uma delas é o limite de velocidade na adaptação. Fiz a hipótese de que a forma
da resposta a cada fóton reflete esta reação que controla a sensibilidade. É,
portanto, importante definir a forma para a resposta a cada fóton. Escolhi a
exponencial simples. Isto é, seguindo uma isomerização, a fotocorrente tem um
aumento rápido seguido por uma queda exponencial, caracterizada por uma
constante de tempo. Esta não é uma má aproximação, desde que os cones e
bastonetes, quando adaptados ao escuro, apresentam este mesmo comportamento [5].
Para analisar um sistema
compartimentalizado, faço uso de um teorema da Mecânica Estatística chamado
"a hipótese ergódica", ou "o teorema da ergodicidade" que
afirma: Para um processo randômico estacionário, um grande número de
observações feitas num único sistema em N instantes arbitrários de tempo tem as
mesmas propriedades estatísticas que observar N sistemas escolhidos
arbitrariamente ao mesmo tempo a partir de um conjunto de sistemas semelhantes.
Igualando compartimento a sistema, posso analisar o comportamento de um único
compartimento através do tempo, sub uma dada condição de luz ambiente, e
predizer o comportamento do fotoreceptor inteiro, formado de um dado número de
compartimentos.
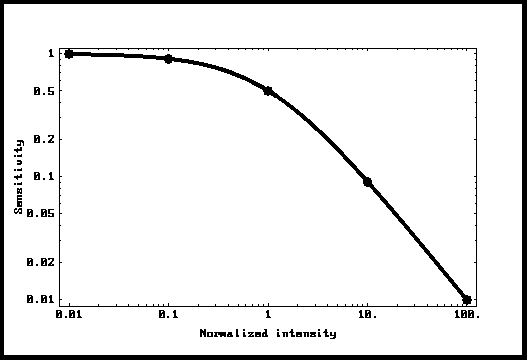
Depois de uma isomerização que
produz uma resposta máxima (normalizada aqui para 1) decaindo exponencialmente
com uma constante de tempo "T", a amplitude da resposta ao fóton
seguinte vai depender do tempo transcorrido desde a isomerização precedente e
será igual a " 1 - Exp[-t/T] ". Como os intervalos entre
isomerizações seguem a distribuição de Poisson, podemos simular uma dada
intensidade para a luz ambiente e encontrar a resposta (média) no
compartimento. Isto foi feito usando Mathematica, e os pontos na Fig.4 são os
resultados desta simulação. Pode-se observar que os pontos caem exatamente
sobre a curva Io/(Io + I). A conclusão é que tal sistema se adapta exatamente
como a lei de Weber-Fechner prediz. Esta é a contribuição maior deste
trabalho.
Figura 4: Sensibilidade como função da
luz ambiente.
A curva de Weber-Fechner.
O parâmetro simples Io, que
até agora foi simplesmente ajustado para acertar os dados experimentais, é
agora associado a estrutura do segmento externo, que é o número de
compartimentos, e a constante de tempo de queda da resposta de cada fóton. A
relação é que Io é gual a N/T, onde N é o número de compartimentos, e T a
constante de tempo de queda.
Os resultados são que, a
partir do conhecimento do tamanho dos compartimentos, o seu número e a
constante de tempo para a resposta a cada fóton, pode-se localizar de forma
absoluta a posição das curvas de intensidade e adaptação no eixo de
intensidade.
Resultados e Discussão
O caso do cone
A pequena resposta a um
flash de um cone mamífero apresenta um pico a mais ou menos 50 mseg.; as
proteínas iodopsina, transducina e fosfodiesterase tem coeficiente de difusão
indo de 0.5 a 2 mícrons quadrados por segundo. Difundindo em duas dimensões,
muito poucas destas moléculas excitadas podem deixar um disco que tem um
diâmetro de 1.5 a 2 microns neste curto período. GMP-cíclico, que difunde mais
rápido, para ser hidrolisado no disco ativo, tem que primeiro encontrar a saída
de seu próprie disco e então encontrar a entrada para o disco ativa. Minha
hipótese é que este é um evento raro. Dentro do disco ativo, pode-se mostrar
por cálculo simples que o número de moléculas ativas de fosfodiesterase pode
ultrapassar o número de moléculas de GMP-c livres presentes. Portanto, a
possibilidade de que todos os canais catiônicos estejam fechados depois de uma
única isomerização existe.
O primeiro resultado desta
análise é que, se dentro dos cones o disco representa um compartimento, e a
resposta dentro daquele compartimento é máxima, então a resposta a um fóton é
1/N, onde N é o número de discos. A partir de fotos de microscopia eletrônica,
sabe-se que o número de discos por micrômetro é por volta de 33. Então a partir
do comprimento total do cone em micrômetros multiplicado por 33 temos o número
de discos. Para o cone da tartaruga, o número de discos é perto de 500, e a
resposta a cada fóton é, então, de 0,2% de mudança, o que está de acordo com os
valores observados experimentalmente de 0,16% [6]. Dados obtidos em 2006[7]oferecem um apoio
adicional ao model 1/N. Em ratos alterados geneticamente pode-se obter cones com
segmentos externos de comprimento variável. Nos ratos Gt alfa -/- os cones S e M
têm segmentos externos de um comprimento de 13.4 mícrons e têm-nos respectivamente
do resposta a um fotão de 0.2 e 0.22 % da resposta máxima. O model 1/N prediz
1/(13.4 x 33) ou 0.23 %. Nos ratos Nrl -/- os cones S têm um segmento externo
de 7.1 mícrons e a resposta a um fotão é 0.44 %, o model prediz 1/(7.1 x 33)
ou 0.43 %. É difícil pedir um melhor acordo entre dados experimentais e um
modelo teórico.Isto também indica porque cones são em
geral curtos; aumentando um disco ao segmento externo do cone aumenta também a
probabilidade de "pegar" um fóton, e reduz ao mesmo tempo a resposta
de 1/N para 1/(N+1). A natureza teve que fazer um compromisso entre pegar
fótons e fornecer uma resposta de tamanho adequado.
A partir das dimensões do
compartimento, ou seja, do disco do cone, é possível calcular um valor para o
fator "k" pela forma de saturação exponencial. [8]. O conhecimento da densidade óptica por
unidade de comprimento e a eficiência quântica são necessários para derivar que
o "k" para cones é igual a: 0,00059 vezes o quadrado do diâmetro do
segmento externo do cone expresso em micrômetros. Esta fórmula foi derivada
para cones estimulados transversalmente; isto corresponde à situação
experimental em que se registra no segmento externo. Ela é válida também para
estimulação axial, que corresponde à situação fisiológica, desde que os
segmentos externos não sejam longos demais. Para cones de macacos, que têm
diâmetro da ordem de 1,5 micrômetros, o valor para "k" calculado com
a fórmula acima é igual a 0,001 micrômetros quadrados, que é próximo ao valor
experimental encontrado de 0,0004 [9].
Para obter uma estimativa
da intensidade de luz ambiente (medida em isomerizações por segundo) que vai
reduzir a sensibilidade em 50%, deve-se simplesmente dividir o número de
compartimentos (800 no caso do cone de tartaruga) pelo valor da constante de
tempo de queda da resposta do cone ao flash tirado dos dados experimentais.
Isto fornece uma taxa de isomerização de 7300 por seg. para Io, enquanto os
valores experimentais estão entre 2000 e 16000 por seg. [10].
O caso dos bastonetes
Para os bastonetes, o
compartimento não é definido por membranas como é o caso dos cones. Aqui temos
uma combinação dos fatos de que as proteínas envolvidas no processo da
fototransdução têm todas um tempo de vida limitado e que a molécula de
GMP-cíclico controlando a abertura dos canais catiônicos tem um coeficiente de
difusão muito mais reduzido no ambiente do segmento externo dos bastonetes. O
tempo para a resposta do bastonete do mamífero atingir o pico é 100 mseg., o
que faz com que mais transducina e mais fosfodiesterase possa ser ativada por
uma única rodopsina. A ação da bioquímica pode se alastrar além do espaço
inter-discos, e estender-se longitudinalmente. A geometria de pilha de discos
no bastonete é também importante no sentido que as moléculas de GMP-cíclico
localizadas entre os discos precisam ser hidrolisadas primeiro se a redução na
concentração deve proceder longitudinalmente. Isso limita a ação da bioquímica,
e estamos na presença do que eu chamo de um "compartimento
operacional".
A solução exata para a
concentração longitudinal de cGMP perto da membrana plasmática, depois de uma
isomerização em um dado disco requereria cálculos complexos em dinâmica de
fluidos. Portanto, obteve-se uma fórmula empírica a partir da análise da
resposta a cada fóton em várias espécies animais. A fórmula diz que os canais
fechados cobrem uma área de membrana plasmática igual àquela contida num disco
(duas folhas) do segmento externo. Por cálculos simples obtemos para o
"comprimento de oclusão" um comprimento do segmento externo igual à
metade do diâmetro. Para o bastonete no sapo e mamíferos isto dá uma redução na
corrente perto de 5%, resultado de uma única isomerização, que é o valor obtido
experimentalmente. Um conjunto de equações semelhante àquele feito para o cone fornece
um valor de "k" para bastonetes igual a 0,01 vezes o cubo do diâmetro
expresso em micrômetros. Pode-se então localizar em termos absolutos a curva
intensidade/resposta no eixo de intensidade. Para os bastonetes dos mamíferos,
o valor calculado para k é 0,08 micrômetros quadrados, enquanto os valores
obtidos experimentalmente são da ordem de 0,006 a 0,07. [8,11]
O número de compartimentos
"N" em bastonetes é obtido dividindo-se o comprimento do segmento
externo por d/2, o que nos fornece um número perto de 20. O parâmetro Io na
relação Weber-Fechner é obtido dividindo 20 pela constante de tempo da
exponencial ajustada à queda da resposta a um fóton. Para os bastonetes de
macacos e humanos, as predições do modelo são que Io deveria estar por volta de
110-113 isomerizações por segundo, enquanto os valores experimentais estão por
volta de 100-120 por segundo. [7,10]
Conclusão
A análise da estrutura geométrica
do segmento externo dos vertebrados fornece resultados interessantes.
o
Primeiramente,
permitiu uma previsão da resposta de cada fóton tanto nos bastonetes quanto nos
cones, e um valor para o parâmetro "k" na fórmula de saturação
exponencial, permitindo uma localização absoluta sobre a curva no eixo de
intensidade.
o
Segundo,
com a ajuda do "teorema de ergodicidade ", pude prever o
comportamento na adaptação de um sistema compartimentalizado. Se a queda da
resposta pode ser aproximada por uma queda exponencial, o resultado é a lei de
"Weber-Fechner" Novamente, esta curva pode ser posicionada no eixo de
intensidade de uma forma absoluta.
Finalmente, o modelo pode
ser testado no sentido em que, em vertebrados de sangue frio, a constante de
tempo da queda da resposta a um fóton muda com a temperatura [12] . Pode-se testar se as propriedades de
adaptação (Io) variam de acordo com isso.
Seus comentários, críticas
e sugestães, serão apreciados. Escreva para: jean-pierre.raynauld@umontreal.ca
1. Lamb
TD, Pugh EN (1992) G-protein cascades: gain and kinetics . Trends Neurosci 15:
291-298.
2. Alfieri
R, Sole P, Gentou C, Kantelip B, Kantelip J-P . (1984) Les cellules
rétiniennes. Ed. D.G.P.L. Paris.
3.
Eckmiller MS (1987) Cone Outer Segment Morphogenesis: Taper Change and Distal
Invaginations. J Cell Biol 105: 2267-2277.
4. Lamb
TD, McNaughton PA, Yau KW (1981) Spatial spread of activation and background
desensitization in toad rod outer segments. J Physiol 319: 463-496.
5. Corson
DW, Cornwall MC, MacNichol EF, Tsang S, Derguini F, Crouch R K,Nakanishi K
(1994) Relief of opsin desensitization and prolonged excitation of rod
photoreceptors by 9-desmethylretinal. Proc Natl Acad Sci USA 91: 6958-6962.
6. Baylor
DA, Hodgkin AL (1973) Detection and resolution of visual stimuli by turtle
photoreceptors. J Physiol 234: 163-198
7. Nikonov SS,
Kholodenko R, Lem J, and Pugh EN. (2006). Physiological features
of the S- and M-cone Photoreceptors of wild-type mice from single-cell recordings.
J.Gen. Physiol. 127(4): 359-374
8. Baylor
DA., Nunn BJ, Schnapf JL (1984) The photocurrent noise and spectral
sensitivities of rods of the monkey Macaca fascicularis. J Physio l 357:
575-607
9. Schnapf
JL, Nunn BJ, Meister M, Baylor DA (1990) Visual transduction in cones of the
monkey Macaca fascicularis. J Physiol 427: 681-713
10. Baylor
DA, Hodgkin AL (1974) Change in time scale and sensitivity in turtle
photoreceptors. J Physiol 242: 729-758.
Burkhardt DA (1994) Light adaptation and photopigment bleaching in cone
photoreceptors in situ in the retina of the turtle. J Neurosci 14(3), 1091
-1105.
11. Kraft
TW, Schneeweis DM, Schnapf JL (1993) Visual Transduction in human
photoreceptors. J Physiol 464: 747-765
12. Lamb
TD (1984) Effects of temperature changes on toad rod photo currents. J Phsysiol
346: 557-578
Do
aquário doméstico de
Jean-Pierre Raynauld
(última atualização em 24 de mayo de 2007)
A versão em Português é uma cortesia de Nathalia L.V. Peixoto